

Structural Insights into the Process of GPCR-G Protein Complex Formation
- PMID: 31080070
- PMCID: PMC6991123
- DOI: 10.1016/j.cell.2019.04.021
Structural Insights into the Process of GPCR-G Protein Complex Formation
Abstract
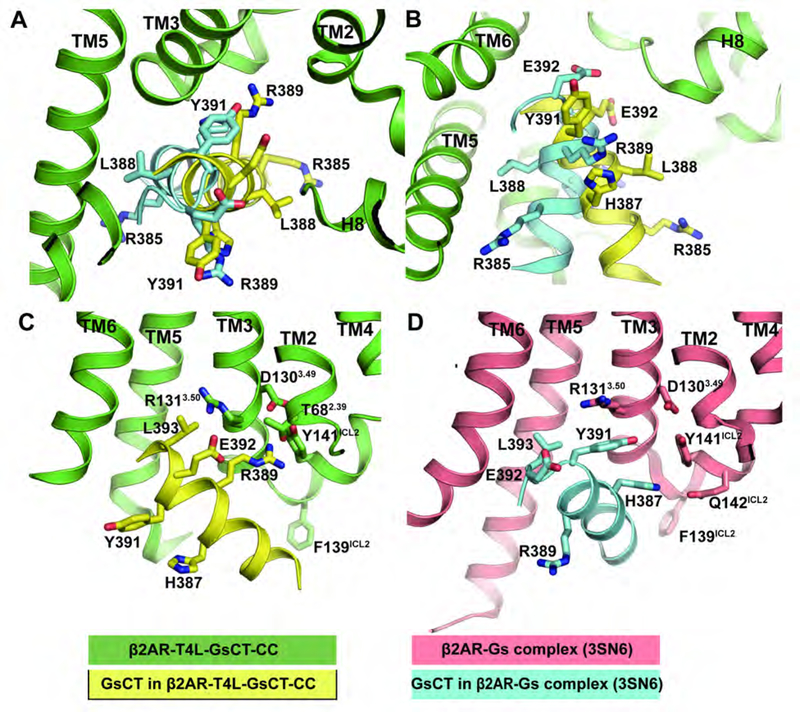
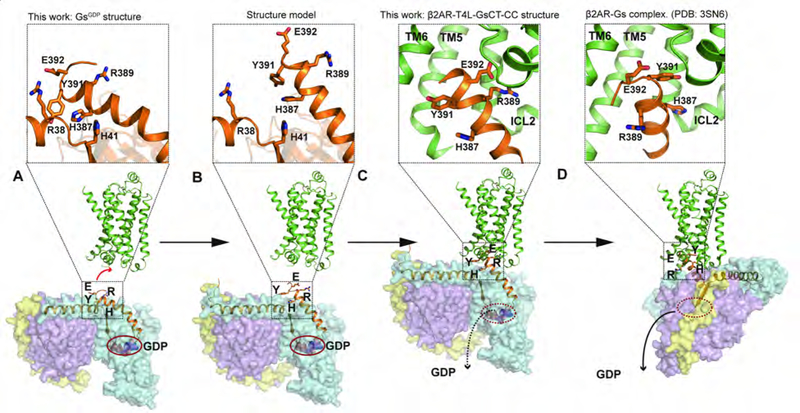
The crystal structure of the β2-adrenergic receptor (β2AR) bound to the G protein adenylyl cyclase stimulatory G protein (Gs) captured the complex in a nucleotide-free state (β2AR-Gsempty). Unfortunately, the β2AR-Gsempty complex does not provide a clear explanation for G protein coupling specificity. Evidence from several sources suggests the existence of a transient complex between the β2AR and GDP-bound Gs protein (β2AR-GsGDP) that may represent an intermediate on the way to the formation of β2AR-Gsempty and may contribute to coupling specificity. Here we present a structure of the β2AR in complex with the carboxyl terminal 14 amino acids from Gαs along with the structure of the GDP-bound Gs heterotrimer. These structures provide evidence for an alternate interaction between the β2AR and Gs that may represent an intermediate that contributes to Gs coupling specificity.
Keywords: G protein; G protein-coupled receptor; coupling specificity; intermediate state; protein engineering.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
B.K.K. is a co-founder of and consultant for ConfometRx, Inc.
Figures





References
-
- Abraham MJM,T; Schulz R; Páll S; Smith JC; Hess B; Lindahl E (2015). GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 1–2, 19–25.
-
- Adams PD, Afonine PV, Bunkoczi G, Chen VB, Davis IW, Echols N, Headd JJ, Hung LW, Kapral GJ, Grosse-Kunstleve RW, et al. (2010). PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta crystallographica Section D, Biological crystallography 66, 213–221. - PMC - PubMed
-
- Baltensperger K, Karoor V, Paul H, Ruoho A, Czech MP, and Malbon CC (1996). The beta-adrenergic receptor is a substrate for the insulin receptor tyrosine kinase. J Biol Chem 271, 1061–1064. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
