

Face-time with TAR: Portraits of an HIV-1 RNA with diverse modes of effector recognition relevant for drug discovery
- PMID: 31080171
- PMCID: PMC6579462
- DOI: 10.1074/jbc.REV119.006860
Face-time with TAR: Portraits of an HIV-1 RNA with diverse modes of effector recognition relevant for drug discovery
Abstract
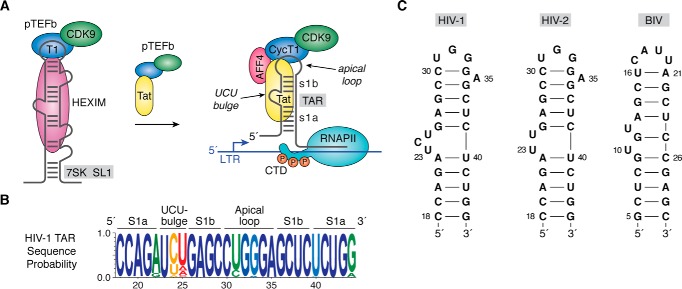
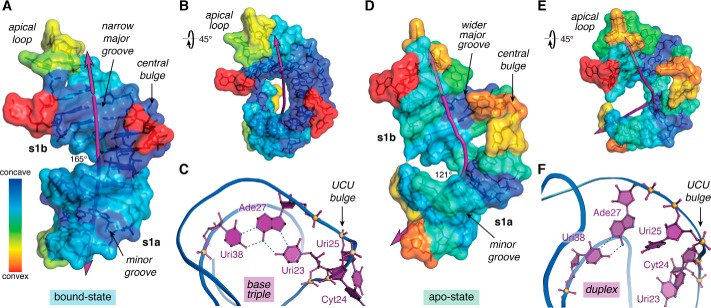
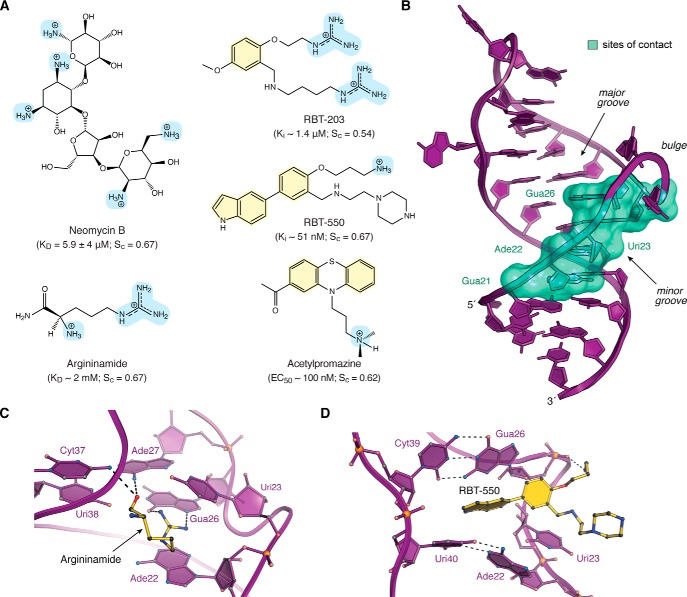
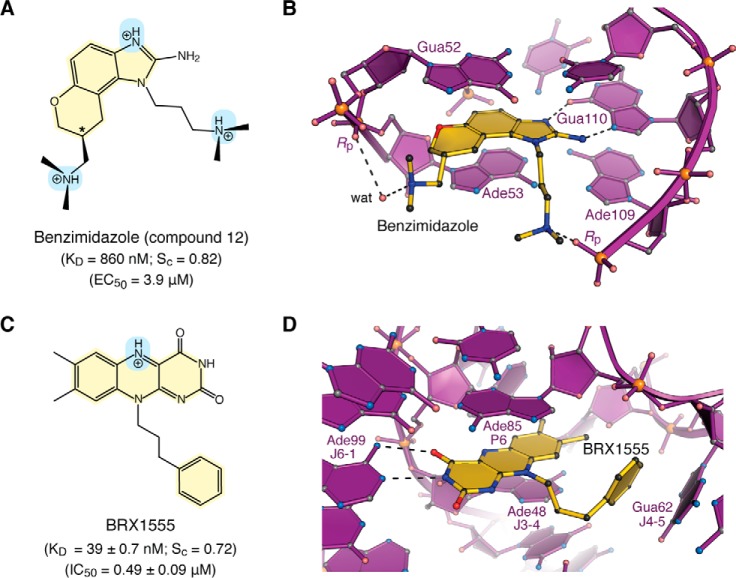
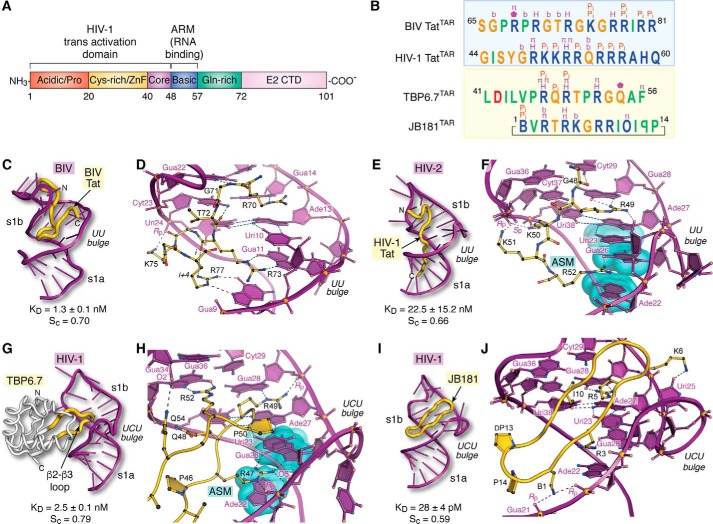
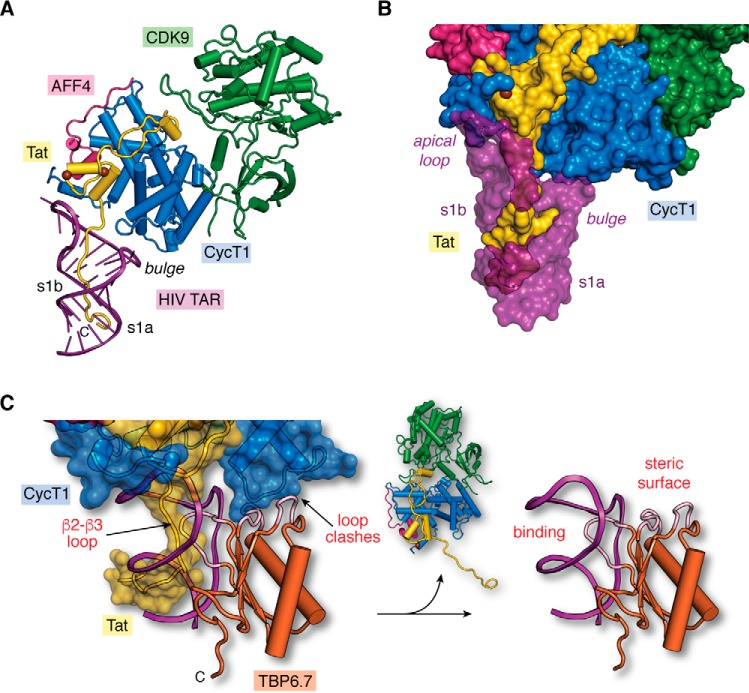
Small molecules and short peptides that potently and selectively bind RNA are rare, making the molecular structures of these complexes highly exceptional. Accordingly, several recent investigations have provided unprecedented structural insights into how peptides and proteins recognize the HIV-1 transactivation response (TAR) element, a 59-nucleotide-long, noncoding RNA segment in the 5' long terminal repeat region of viral transcripts. Here, we offer an integrated perspective on these advances by describing earlier progress on TAR binding to small molecules, and by drawing parallels to recent successes in the identification of compounds that target the hepatitis C virus internal ribosome entry site (IRES) and the flavin-mononucleotide riboswitch. We relate this work to recent progress that pinpoints specific determinants of TAR recognition by: (i) viral Tat proteins, (ii) an innovative lab-evolved TAR-binding protein, and (iii) an ultrahigh-affinity cyclic peptide. New structural details are used to model the TAR-Tat-super-elongation complex (SEC) that is essential for efficient viral transcription and represents a focal point for antiviral drug design. A key prediction is that the Tat transactivation domain makes modest contacts with the TAR apical loop, whereas its arginine-rich motif spans the entire length of the TAR major groove. This expansive interface has significant implications for drug discovery and design, and it further suggests that future lab-evolved proteins could be deployed to discover steric restriction points that block Tat-mediated recruitment of the host SEC to HIV-1 TAR.
Keywords: HIV TAR RNA; RNA structure; RNA virus; RNA-binding protein; RNA–drug interaction; RNA–protein interaction; arginine-sandwich motif; drug development; drug discovery; structural biology; structural model; structure–function; super-elongation complex; viral transactivation.
© 2019 Chavali et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
