

Recent Advances in Understanding σ70-Dependent Transcription Initiation Mechanisms
- PMID: 31082441
- PMCID: PMC7057261
- DOI: 10.1016/j.jmb.2019.04.046
Recent Advances in Understanding σ70-Dependent Transcription Initiation Mechanisms
Abstract
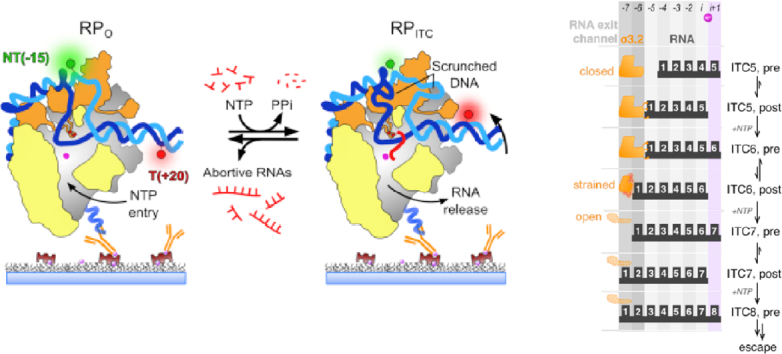
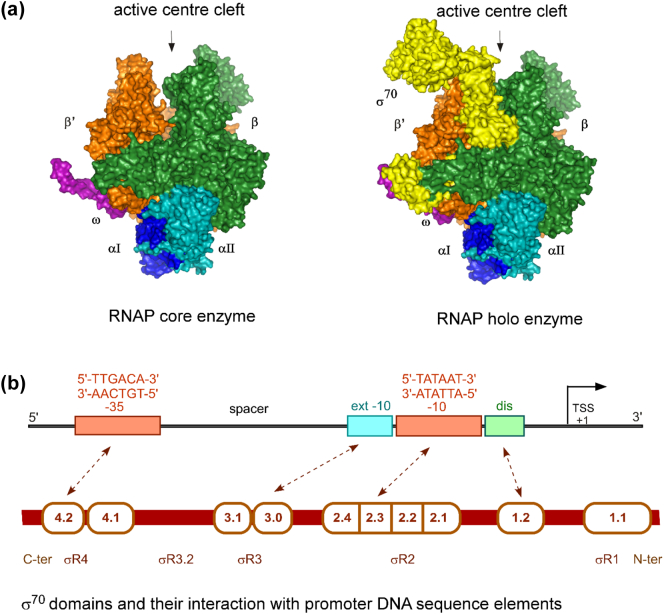
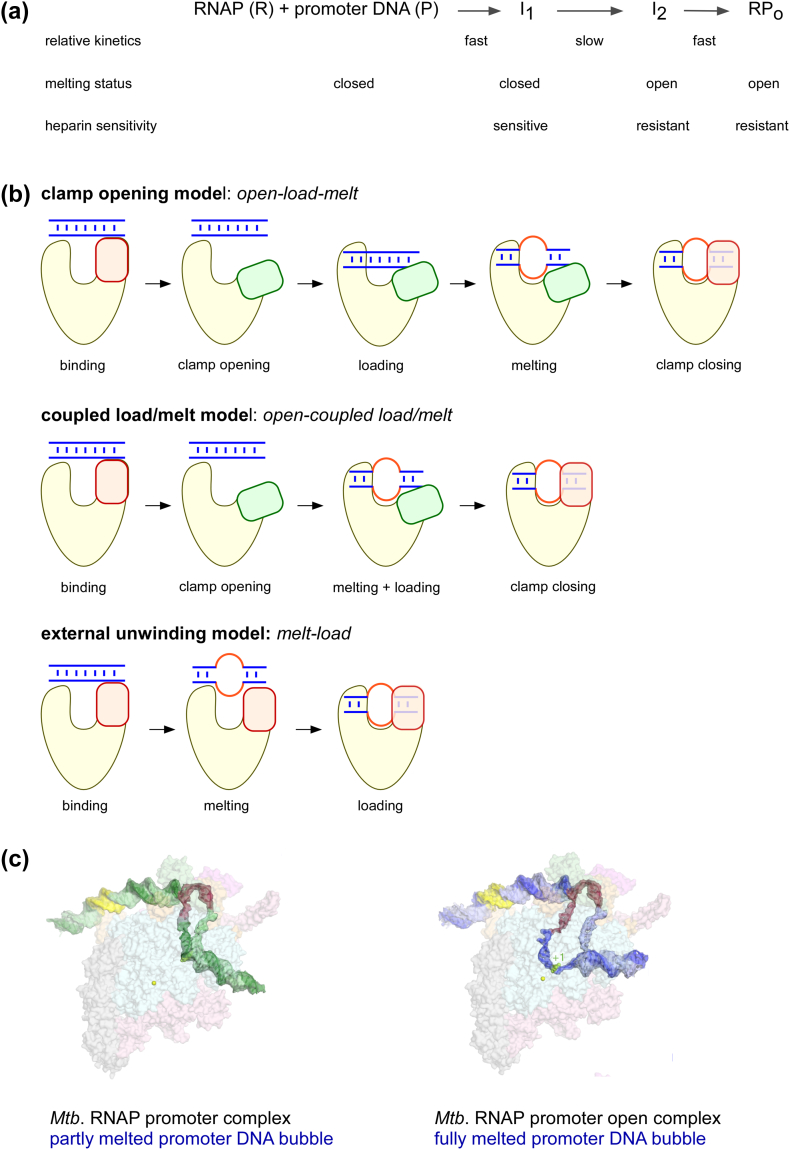
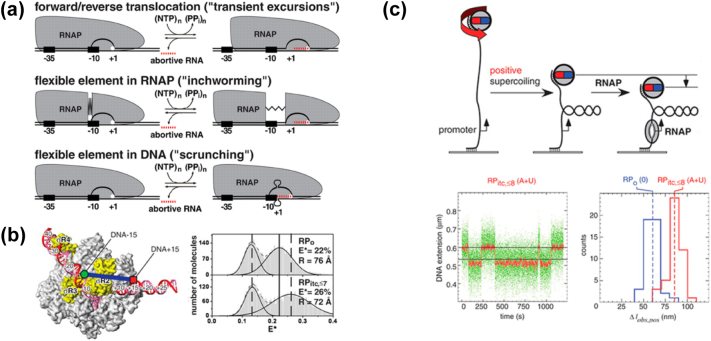
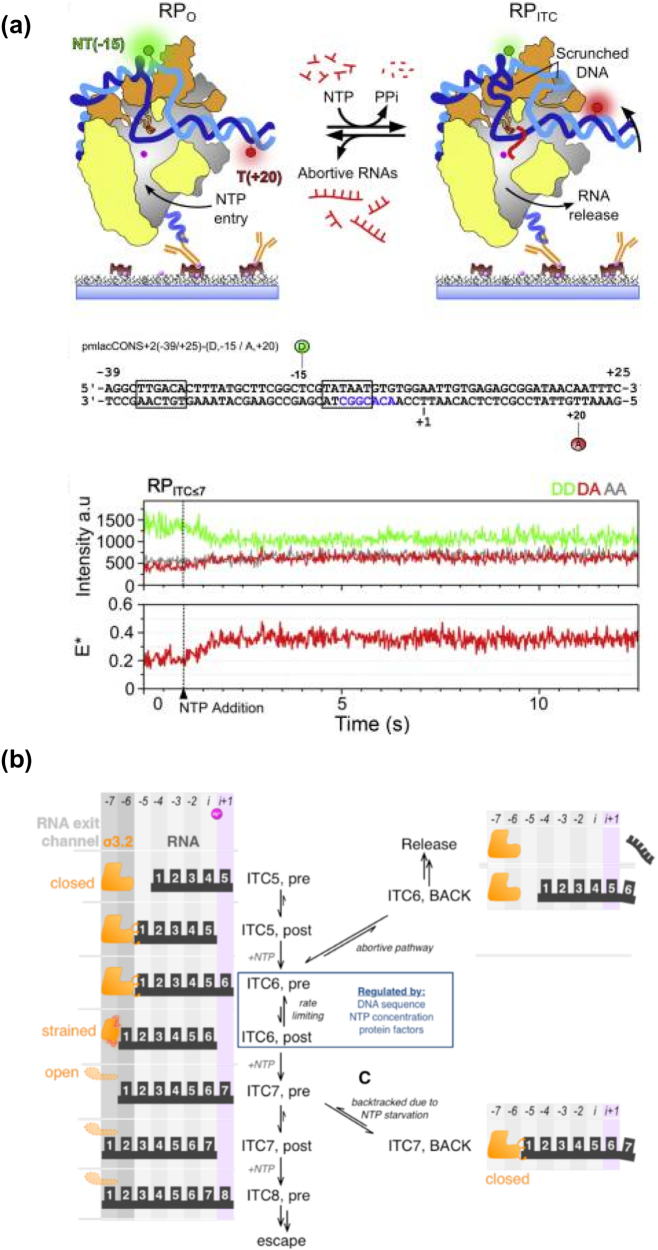
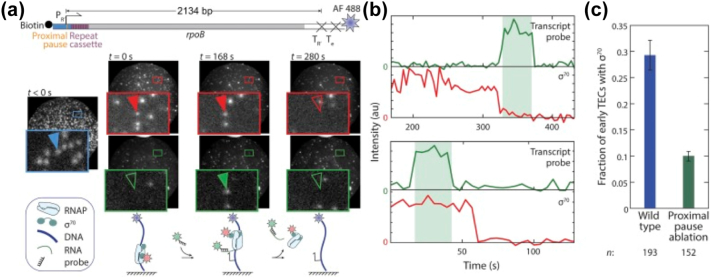
Prokaryotic transcription is one of the most studied biological systems, with relevance to many fields including the development and use of antibiotics, the construction of synthetic gene networks, and the development of many cutting-edge methodologies. Here, we discuss recent structural, biochemical, and single-molecule biophysical studies targeting the mechanisms of transcription initiation in bacteria, including the formation of the open complex, the reaction of initial transcription, and the promoter escape step that leads to elongation. We specifically focus on the mechanisms employed by the RNA polymerase holoenzyme with the housekeeping sigma factor σ70. The recent progress provides answers to long-held questions, identifies intriguing new behaviours, and opens up fresh questions for the field of transcription.
Copyright © 2019 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Decker K.B., Hinton D.M. Transcription regulation at the core: similarities among bacterial, archaeal, and eukaryotic RNA polymerases. Annu. Rev. Microbiol. 2013;67:113–139. - PubMed
-
- Dickson R.C., Abelson J., Barnes W.M., Reznikoff W.S. Genetic regulation: the lac control region. Science. 1975;187:27–35. - PubMed
-
- Pribnow D. Bacteriophage T7 early promoters: nucleotide sequences of two RNA polymerase binding sites. J. Mol. Biol. 1975;99:419–443. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
