

Causes and Biophysical Consequences of Cellulose Production by Pseudomonas fluorescens SBW25 at the Air-Liquid Interface
- PMID: 31085696
- PMCID: PMC6707908
- DOI: 10.1128/JB.00110-19
Causes and Biophysical Consequences of Cellulose Production by Pseudomonas fluorescens SBW25 at the Air-Liquid Interface
Abstract
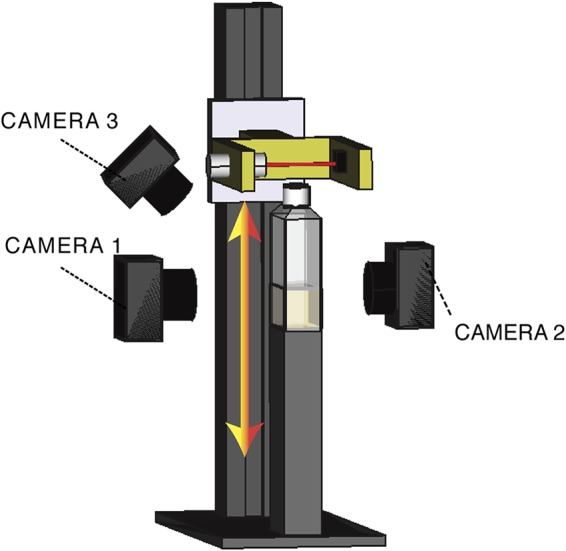
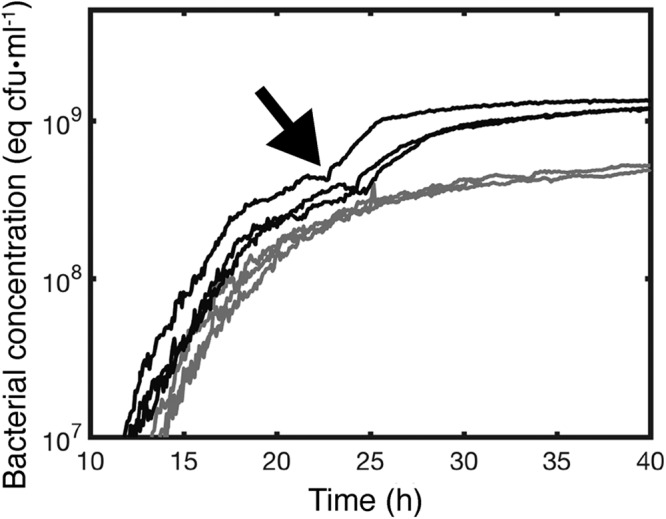
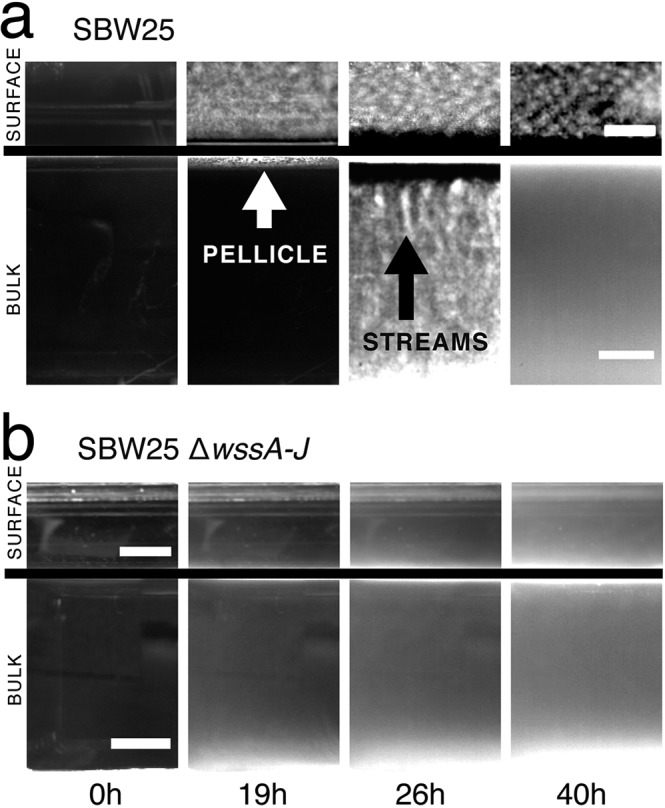
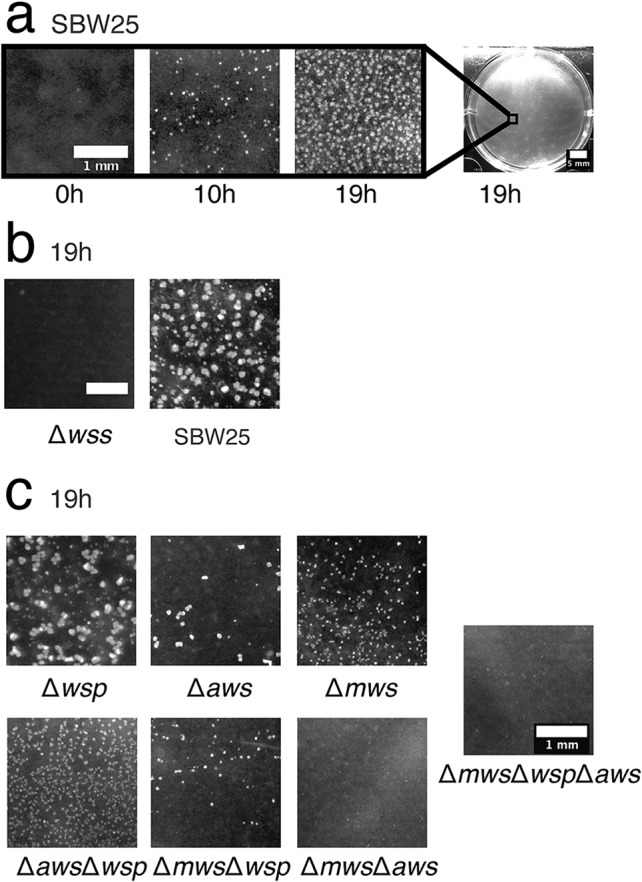
Cellulose-overproducing wrinkly spreader mutants of Pseudomonas fluorescens SBW25 have been the focus of much investigation, but conditions promoting the production of cellulose in ancestral strain SBW25 and its effects and consequences have escaped in-depth investigation through lack of an in vitro phenotype. Here, using a custom-built device, we reveal that in static broth microcosms, ancestral SBW25 encounters environmental signals at the air-liquid interface that activate, via three diguanylate cyclase-encoding pathways (Wsp, Aws, and Mws), production of cellulose. Secretion of the polymer at the meniscus leads to modification of the environment and growth of numerous microcolonies that extend from the surface. Accumulation of cellulose and associated microbial growth leads to Rayleigh-Taylor instability resulting in bioconvection and rapid transport of water-soluble products over tens of millimeters. Drawing upon data, we built a mathematical model that recapitulates experimental results and captures the interactions between biological, chemical and physical processes.IMPORTANCE This work reveals a hitherto unrecognized behavior that manifests at the air-liquid interface that depends on production of cellulose and hints at undiscovered dimensions to bacterial life at surfaces. Additionally, the study links activation of known diguanylate cyclase-encoding pathways to cellulose expression and to signals encountered at the meniscus. Further significance stems from recognition of the consequences of fluid instabilities arising from surface production of cellulose for transport of water-soluble products over large distances.
Keywords: continuum field models; microbial mats; pellicle; pyoverdin; spatial structure.
Copyright © 2019 Ardré et al.
Figures

References
-
- Marshall KC. 1996. Adhesion as a strategy for access to nutrients, p 59–87. In Fletcher M. (ed), Bacterial adhesion: molecular and ecological diversity. Wiley-Liss, Inc, New York, NY.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
