

Islands of retroelements are major components of Drosophila centromeres
- PMID: 31086362
- PMCID: PMC6516634
- DOI: 10.1371/journal.pbio.3000241
Islands of retroelements are major components of Drosophila centromeres
Abstract
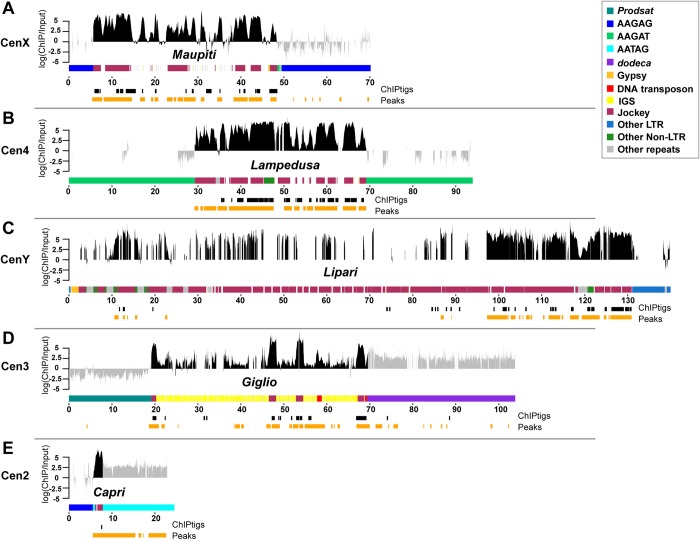
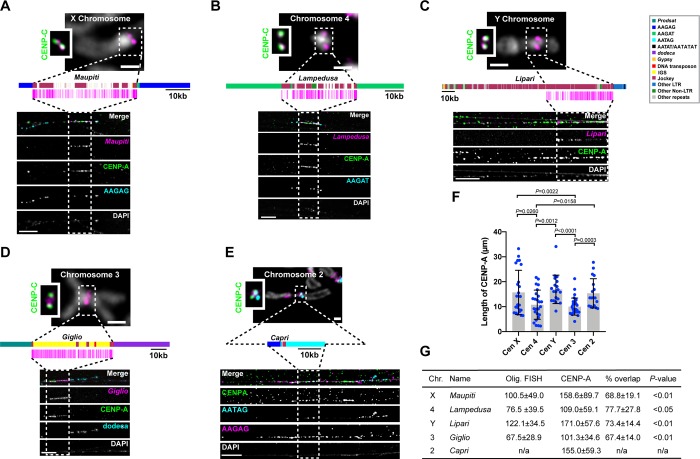
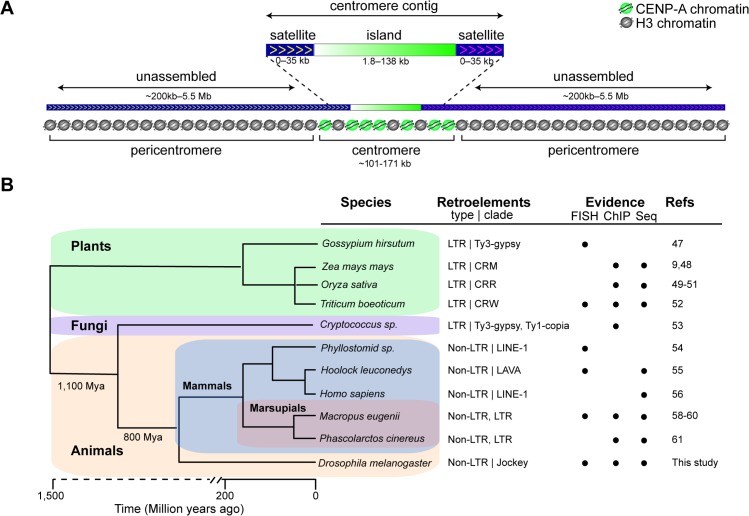
Centromeres are essential chromosomal regions that mediate kinetochore assembly and spindle attachments during cell division. Despite their functional conservation, centromeres are among the most rapidly evolving genomic regions and can shape karyotype evolution and speciation across taxa. Although significant progress has been made in identifying centromere-associated proteins, the highly repetitive centromeres of metazoans have been refractory to DNA sequencing and assembly, leaving large gaps in our understanding of their functional organization and evolution. Here, we identify the sequence composition and organization of the centromeres of Drosophila melanogaster by combining long-read sequencing, chromatin immunoprecipitation for the centromeric histone CENP-A, and high-resolution chromatin fiber imaging. Contrary to previous models that heralded satellite repeats as the major functional components, we demonstrate that functional centromeres form on islands of complex DNA sequences enriched in retroelements that are flanked by large arrays of satellite repeats. Each centromere displays distinct size and arrangement of its DNA elements but is similar in composition overall. We discover that a specific retroelement, G2/Jockey-3, is the most highly enriched sequence in CENP-A chromatin and is the only element shared among all centromeres. G2/Jockey-3 is also associated with CENP-A in the sister species D. simulans, revealing an unexpected conservation despite the reported turnover of centromeric satellite DNA. Our work reveals the DNA sequence identity of the active centromeres of a premier model organism and implicates retroelements as conserved features of centromeric DNA.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
