

IKKβ slows Huntington's disease progression in R6/1 mice
- PMID: 31088970
- PMCID: PMC6561205
- DOI: 10.1073/pnas.1814246116
IKKβ slows Huntington's disease progression in R6/1 mice
Abstract
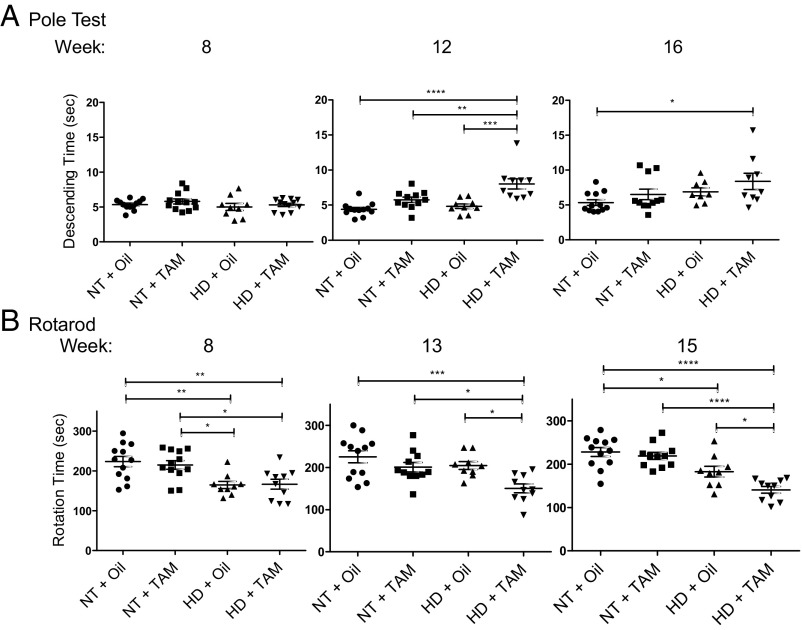
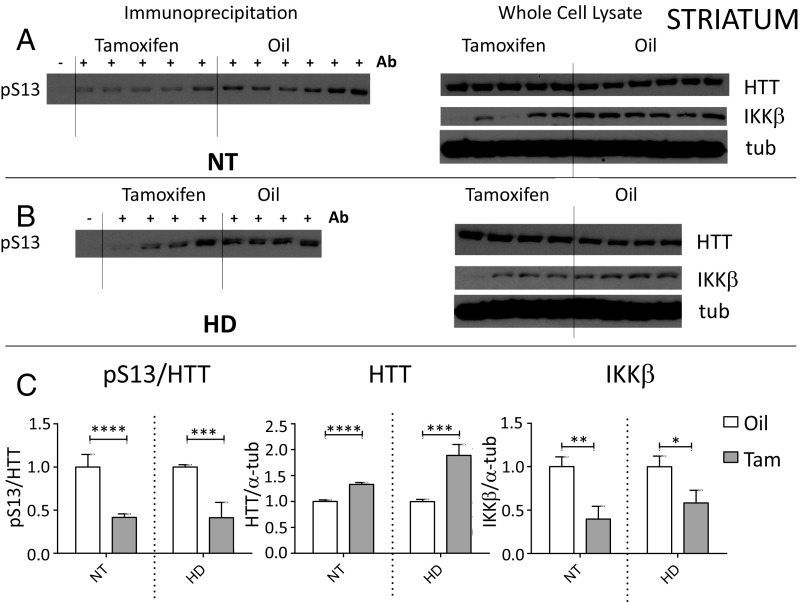
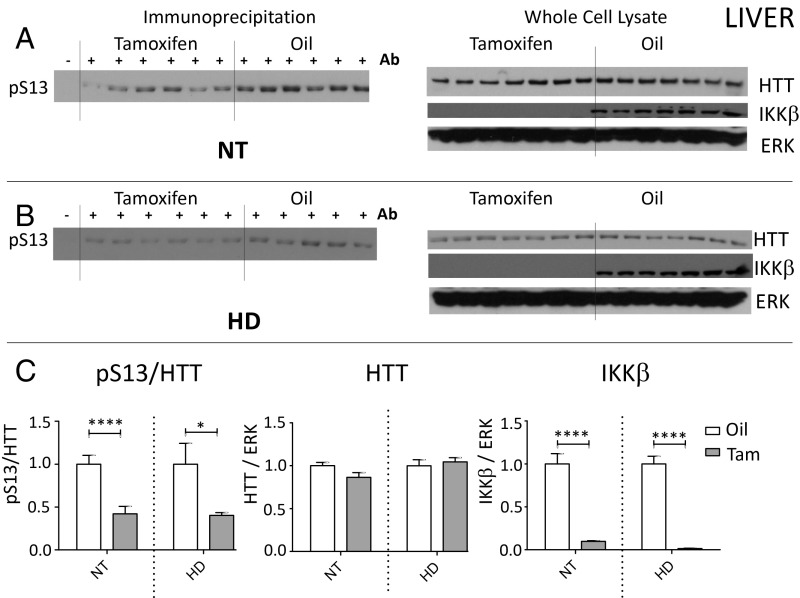
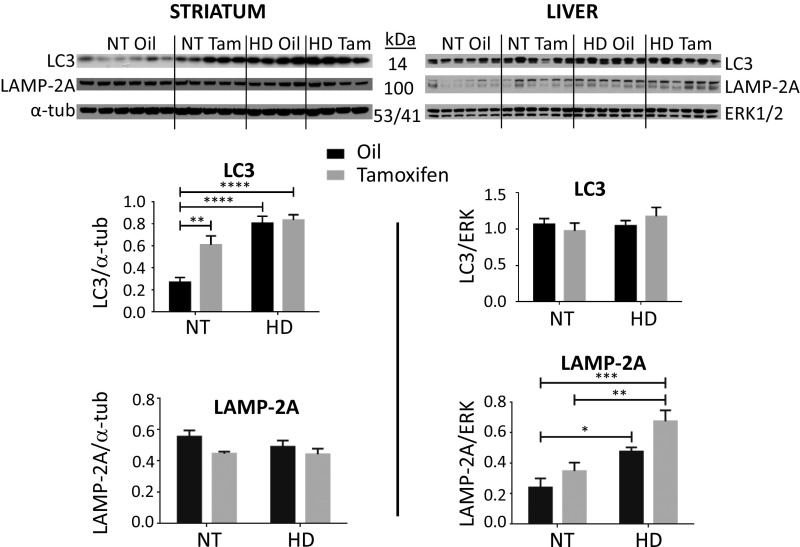
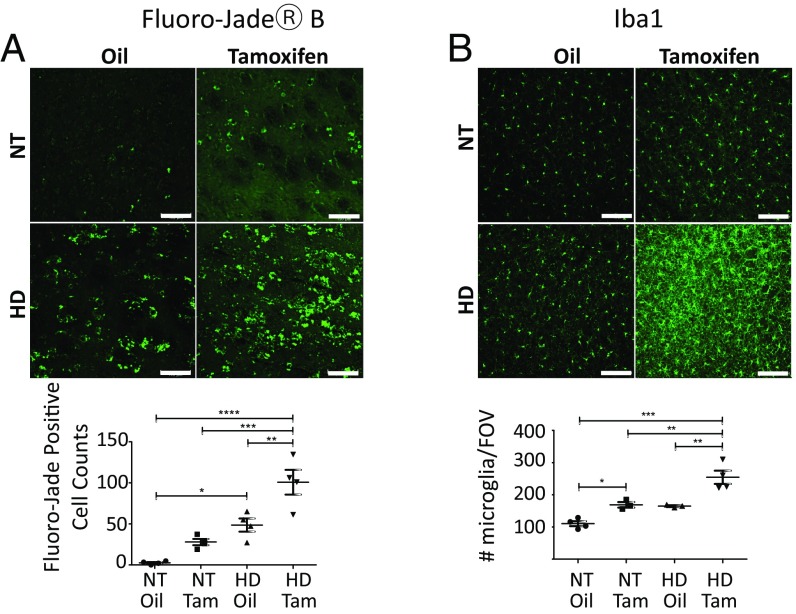
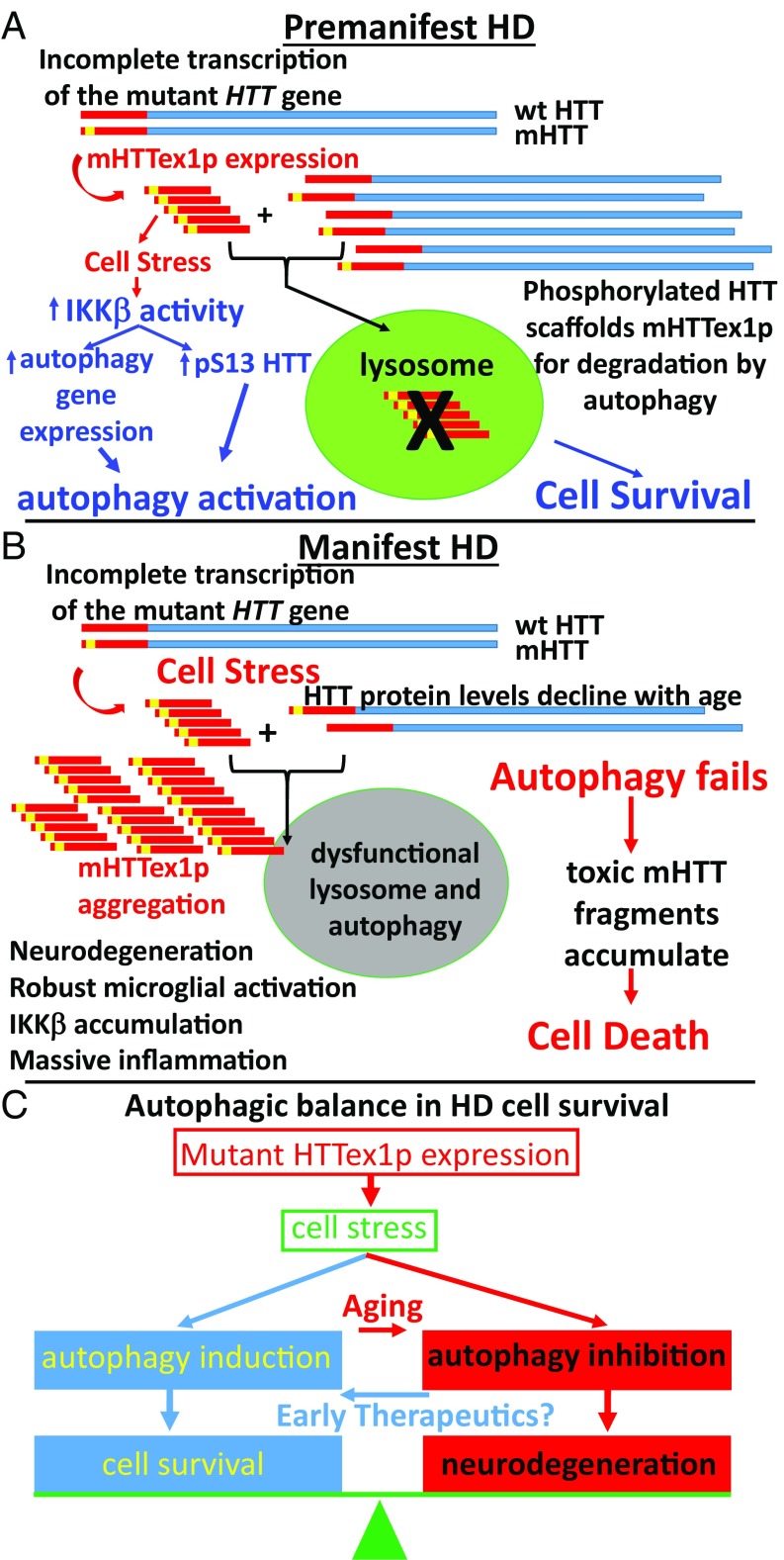
Neuroinflammation is an important contributor to neuronal pathology and death in neurodegenerative diseases and neuronal injury. Therapeutic interventions blocking the activity of the inflammatory kinase IKKβ, a key regulator of neuroinflammatory pathways, is protective in several animal models of neurodegenerative disease and neuronal injury. In Huntington's disease (HD), however, significant questions exist as to the impact of blocking or diminishing the activity of IKKβ on HD pathology given its potential role in Huntingtin (HTT) degradation. In cell culture, IKKβ phosphorylates HTT serine (S) 13 and activates HTT degradation, a process that becomes impaired with polyQ expansion. To investigate the in vivo relationship of IKKβ to HTT S13 phosphorylation and HD progression, we crossed conditional tamoxifen-inducible IKKβ knockout mice with R6/1 HD mice. Behavioral assays in these mice showed a significant worsening of HD pathological phenotypes. The increased behavioral pathology correlated with reduced levels of endogenous mouse full-length phospho-S13 HTT, supporting the importance of IKKβ in the phosphorylation of HTT S13 in vivo. Notably, many striatal autophagy genes were up-regulated in HD vs. control mice; however, IKKβ knockout partially reduced this up-regulation in HD, increased striatal neurodegeneration, and enhanced an activated microglial response. We propose that IKKβ is protective in striatal neurons early in HD progression via phosphorylation of HTT S13. As IKKβ is also required for up-regulation of some autophagy genes and HTT is a scaffold for selective autophagy, IKKβ may influence autophagy through multiple mechanisms to maintain healthy striatal function, thereby reducing neuronal degeneration to slow HD onset.
Keywords: Huntington’s disease; IκB kinase; autophagy; huntingtin; neurodegeneration.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
