

Plant condensin II is required for the correct spatial relationship between centromeres and rDNA arrays
- PMID: 31092096
- PMCID: PMC6527393
- DOI: 10.1080/19491034.2019.1616507
Plant condensin II is required for the correct spatial relationship between centromeres and rDNA arrays
Abstract
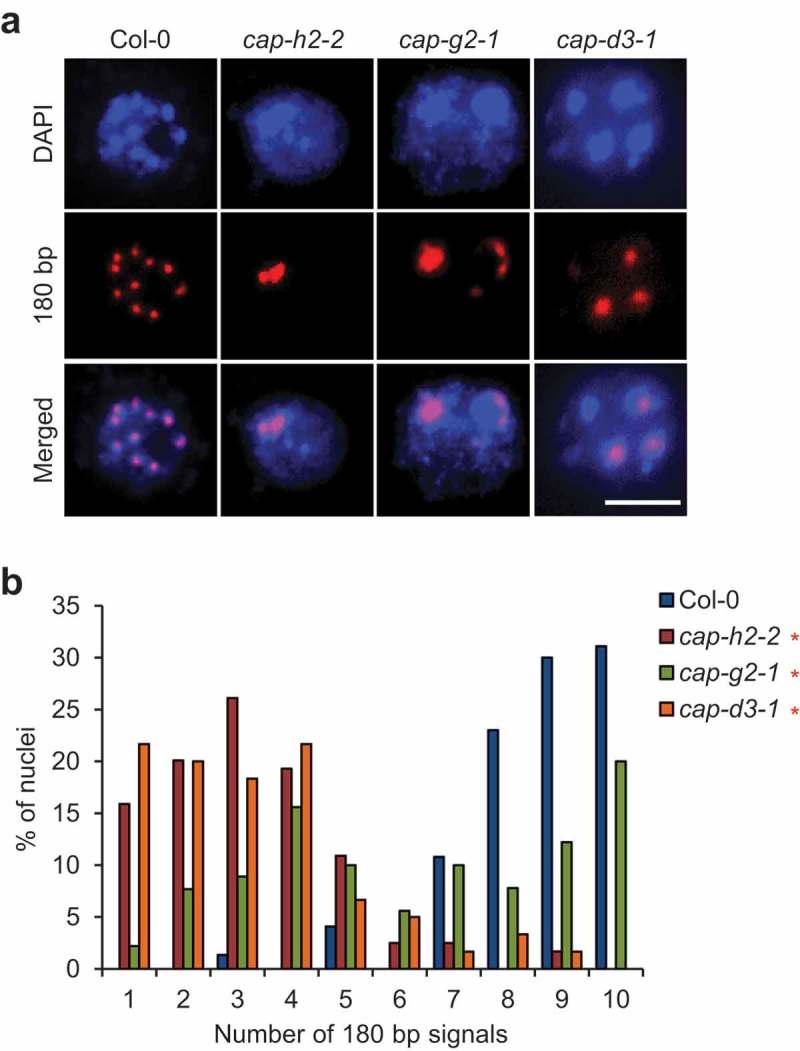
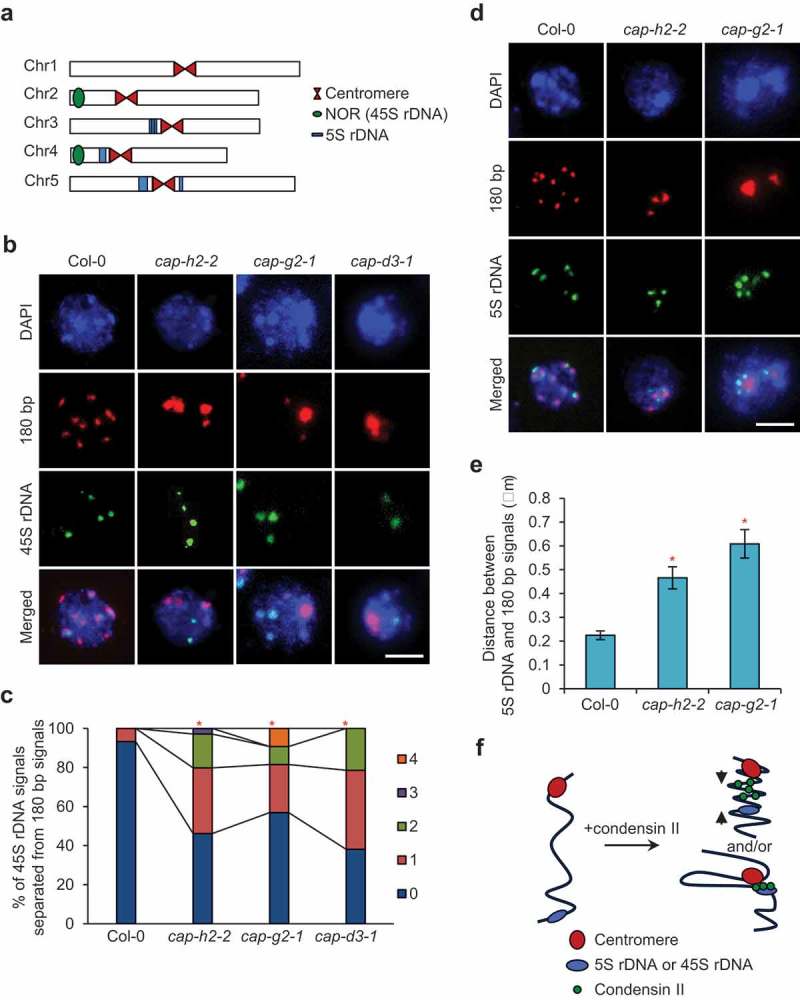
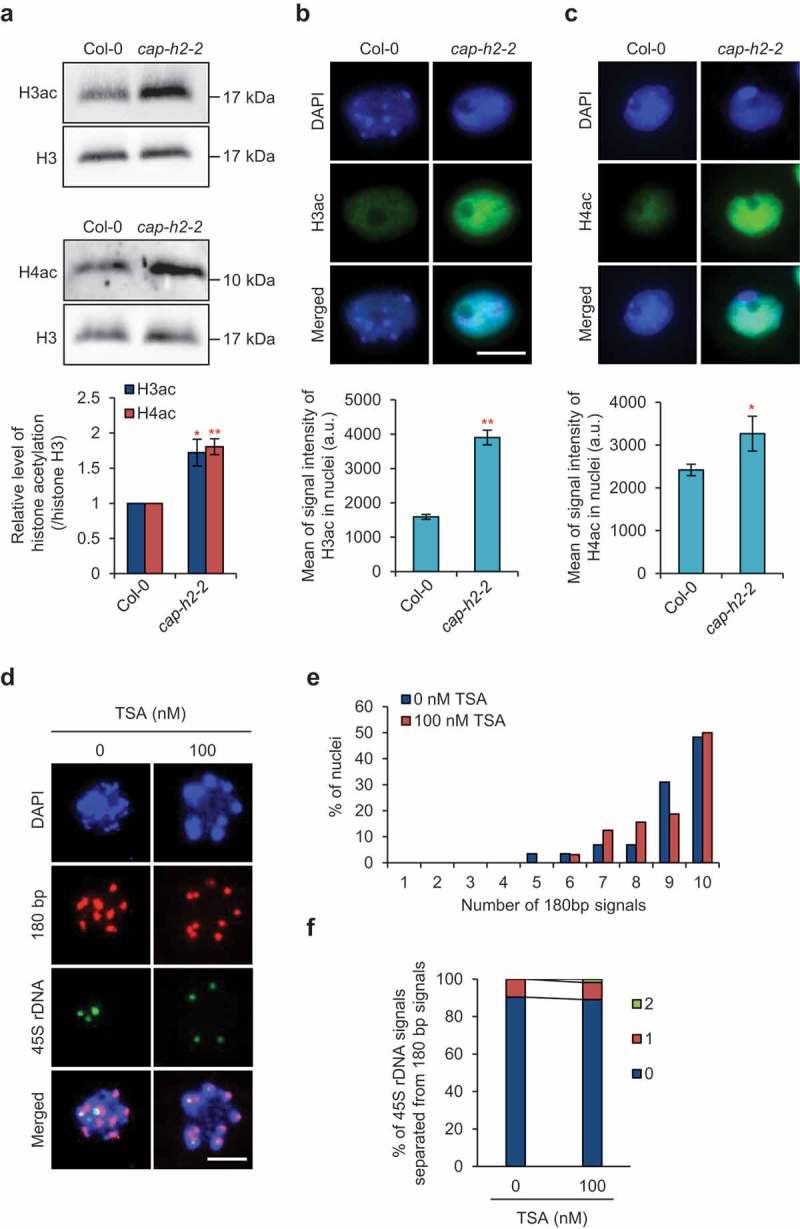
Plants possess the structural maintenance of chromosome (SMC) protein complexes cohesin, condensin, and SMC5/6, which function in fundamental biological processes such as sister chromatid cohesion, chromosome condensation and segregation, and damaged DNA repair. Recently, increasing evidence in several organisms has suggested that condensin is involved in chromatin organizations during interphase. In Arabidopsis thaliana, condensin II is localized in the nucleus throughout interphase and is suggested to be required for keeping centromeres apart and the assembly of euchromatic chromosome arms. However, it remains unclear how condensin II organizes chromatin associations. Here, we first showed the high possibility that the function of condensin II as a complex is required for the disassociation of centromeres. Analysis of the rDNA array distribution revealed that condensin II is also indispensable for the association of centromeres with rDNA arrays. Reduced axial compaction of chromosomes and impaired genome integrity in condensin II mutants are not related to the disruption of chromatin organization. In contrast, the axial compaction of chromosomes by condensin II produces the force leading to the disassociation of heterologous centromeres in Drosophila melanogaster. Taken together, our data imply that the condensin II function in chromatin organization differs among eukaryotes.
Keywords: Condensin II; centromere; genome integrity; histone hyperacetylation; rDNA.
Figures
References
-
- Jeppsson K, Kanno T, Shirahige K, et al. The maintenance of chromosome structure: positioning and functioning of SMC complexes. Nat Rev Mol Cell Biol. 2014;15:601–614. - PubMed
-
- Uhlmann F. SMC complexes: from DNA to chromosomes. Nat Rev Mol Cell Biol. 2016;17:399–412. - PubMed
-
- Hirano T. Condensins: organizing and segregating the genome. Curr Biol. 2005;15:R265–275. - PubMed
-
- Hirano T. Condensin-based chromosome organization from bacteria to vertebrates. Cell. 2016;164:847–857. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases