

Lipid metabolism in cancer cells under metabolic stress
- PMID: 31092908
- PMCID: PMC6738079
- DOI: 10.1038/s41416-019-0451-4
Lipid metabolism in cancer cells under metabolic stress
Abstract
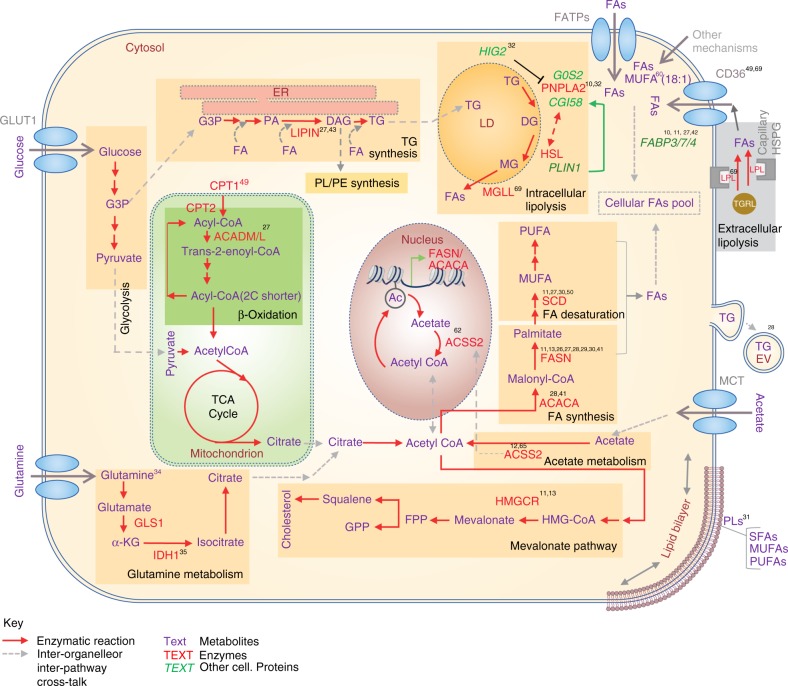
Cancer cells are often exposed to a metabolically challenging environment with scarce availability of oxygen and nutrients. This metabolic stress leads to changes in the balance between the endogenous synthesis and exogenous uptake of fatty acids, which are needed by cells for membrane biogenesis, energy production and protein modification. Alterations in lipid metabolism and, consequently, lipid composition have important therapeutic implications, as they affect the survival, membrane dynamics and therapy response of cancer cells. In this article, we provide an overview of recent insights into the regulation of lipid metabolism in cancer cells under metabolic stress and discuss how this metabolic adaptation helps cancer cells thrive in a harsh tumour microenvironment.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
