

Mitochondrial fragmentation drives selective removal of deleterious mtDNA in the germline
- PMID: 31092924
- PMCID: PMC6614061
- DOI: 10.1038/s41586-019-1213-4
Mitochondrial fragmentation drives selective removal of deleterious mtDNA in the germline
Abstract
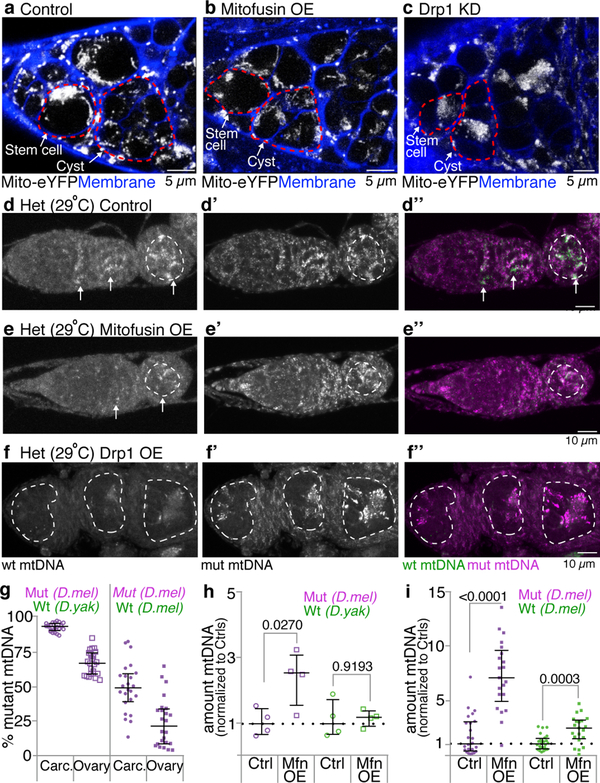
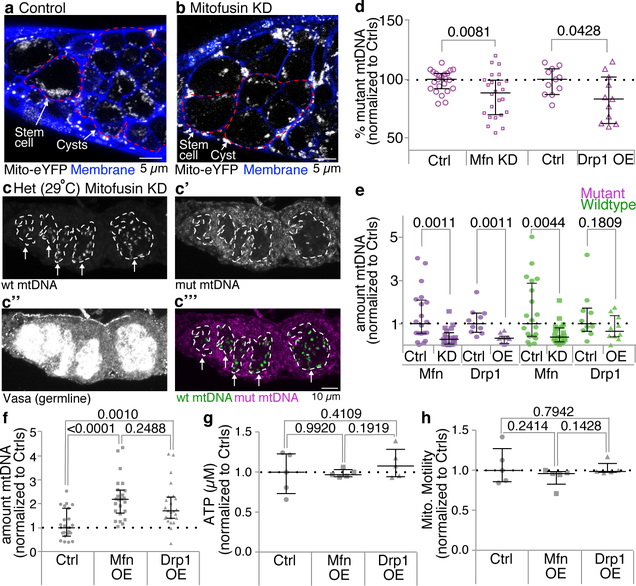
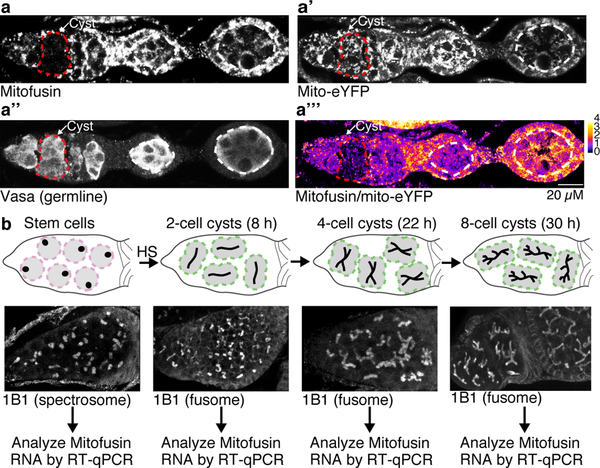
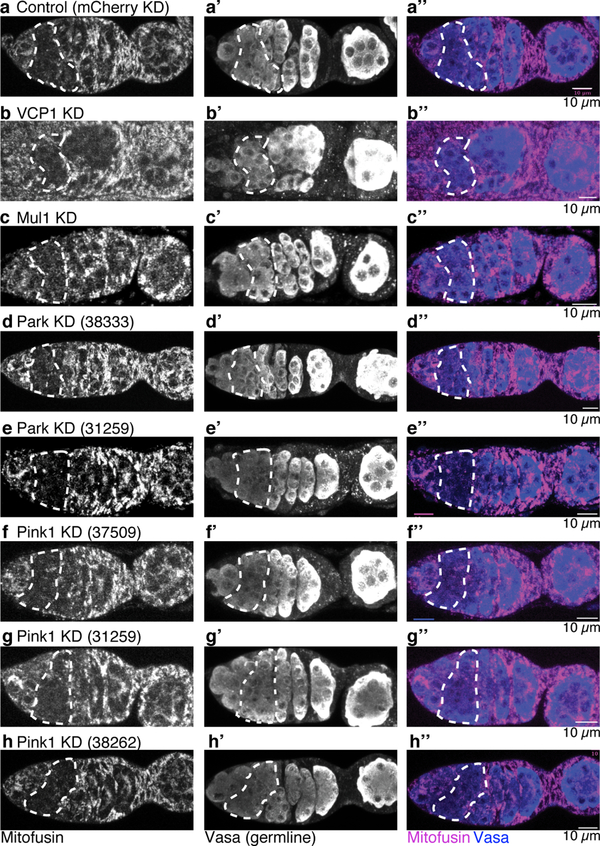
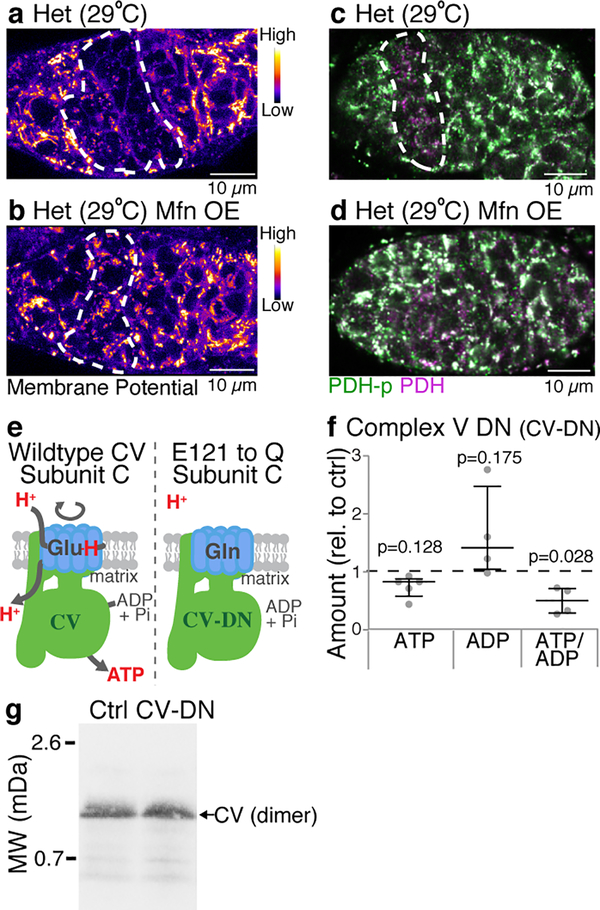
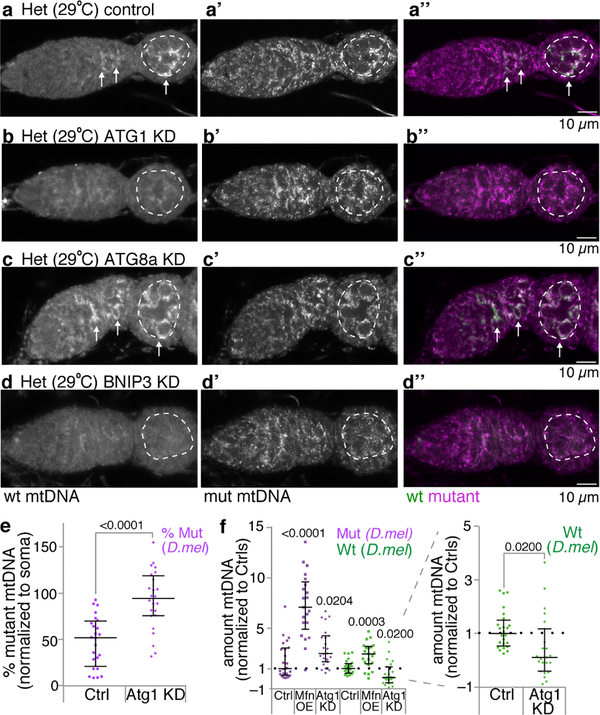
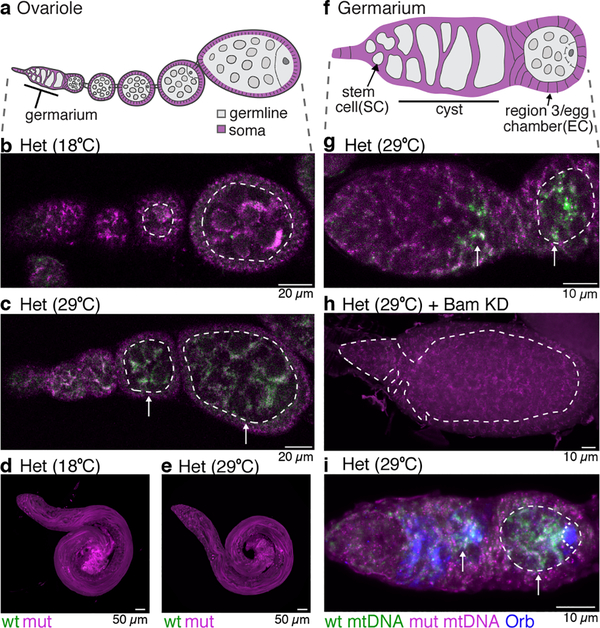
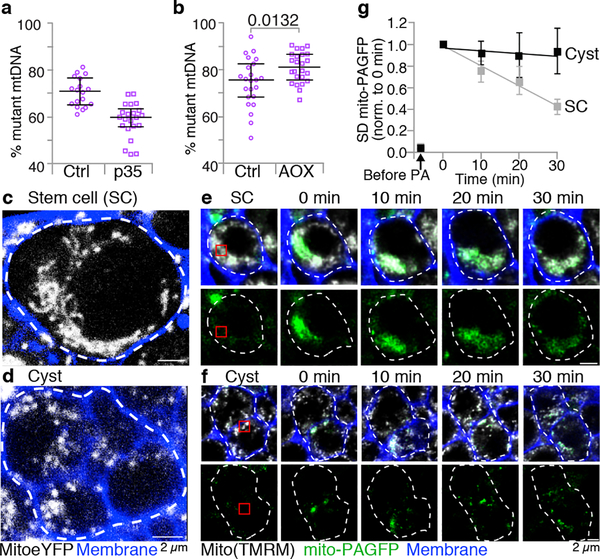
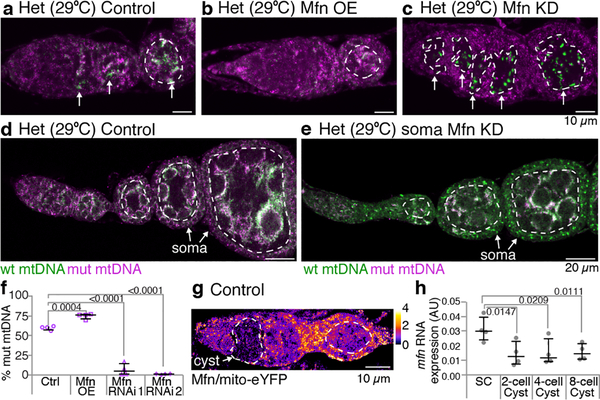
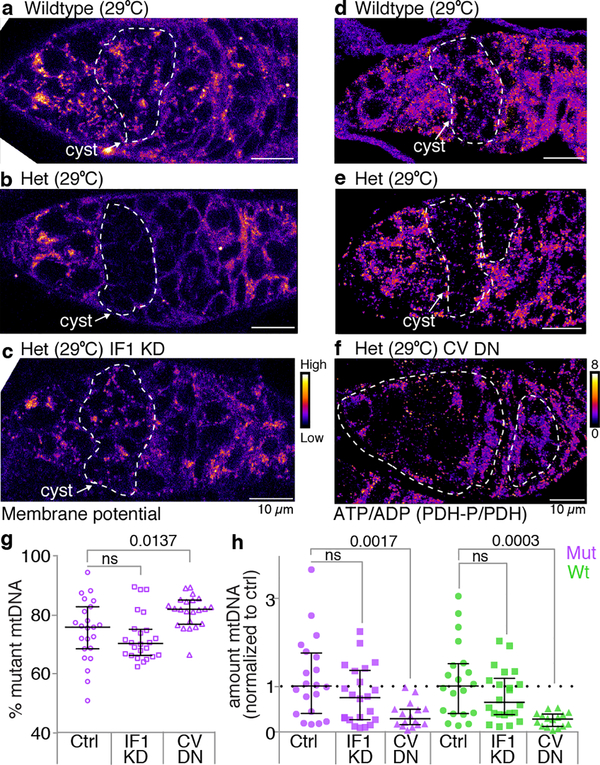
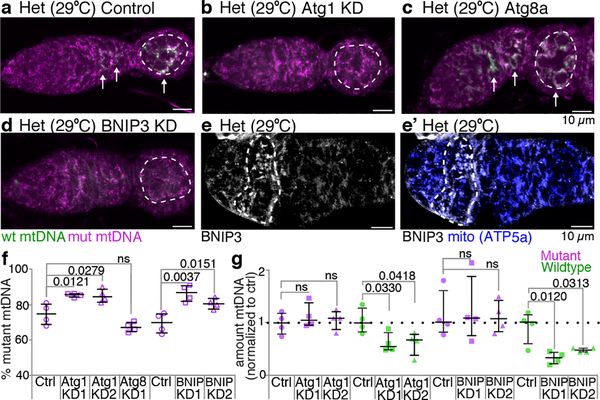
Mitochondria contain their own genomes that, unlike nuclear genomes, are inherited only in the maternal line. Owing to a high mutation rate and low levels of recombination of mitrochondrial DNA (mtDNA), special selection mechanisms exist in the female germline to prevent the accumulation of deleterious mutations1-5. However, the molecular mechanisms that underpin selection are poorly understood6. Here we visualize germline selection in Drosophila using an allele-specific fluorescent in situ-hybridization approach to distinguish wild-type from mutant mtDNA. Selection first manifests in the early stages of Drosophila oogenesis, triggered by reduction of the pro-fusion protein Mitofusin. This leads to the physical separation of mitochondrial genomes into different mitochondrial fragments, which prevents the mixing of genomes and their products and thereby reduces complementation. Once fragmented, mitochondria that contain mutant genomes are less able to produce ATP, which marks them for selection through a process that requires the mitophagy proteins Atg1 and BNIP3. A reduction in Atg1 or BNIP3 decreases the amount of wild-type mtDNA, which suggests a link between mitochondrial turnover and mtDNA replication. Fragmentation is not only necessary for selection in germline tissues, but is also sufficient to induce selection in somatic tissues in which selection is normally absent. We postulate that there is a generalizable mechanism for selection against deleterious mtDNA mutations, which may enable the development of strategies for the treatment of mtDNA disorders.
Figures
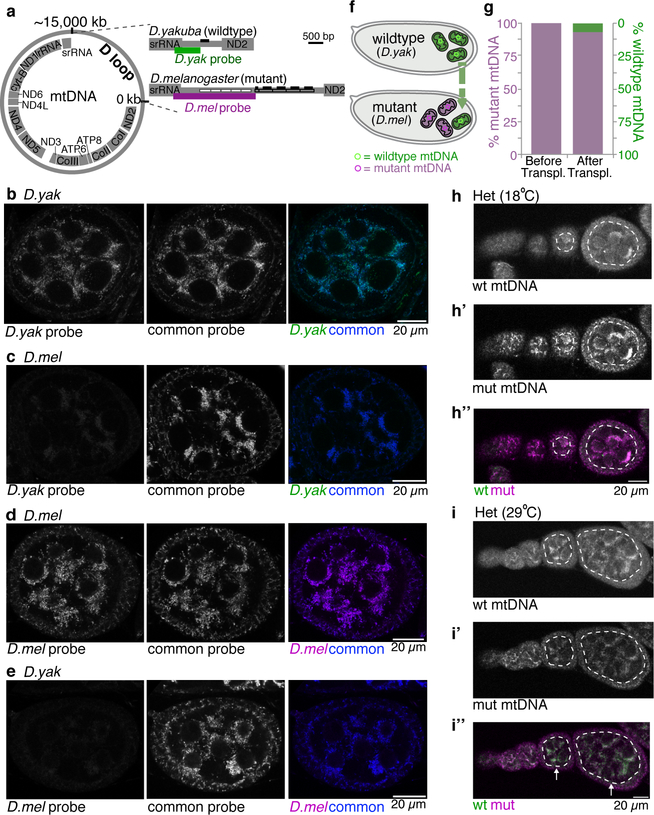
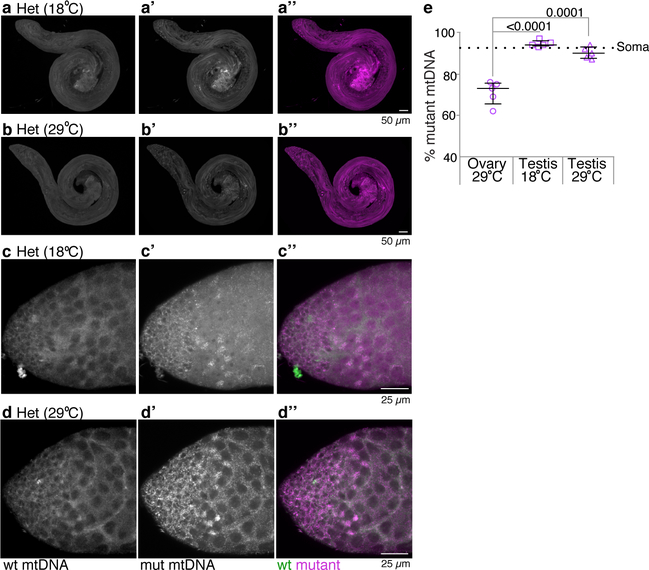
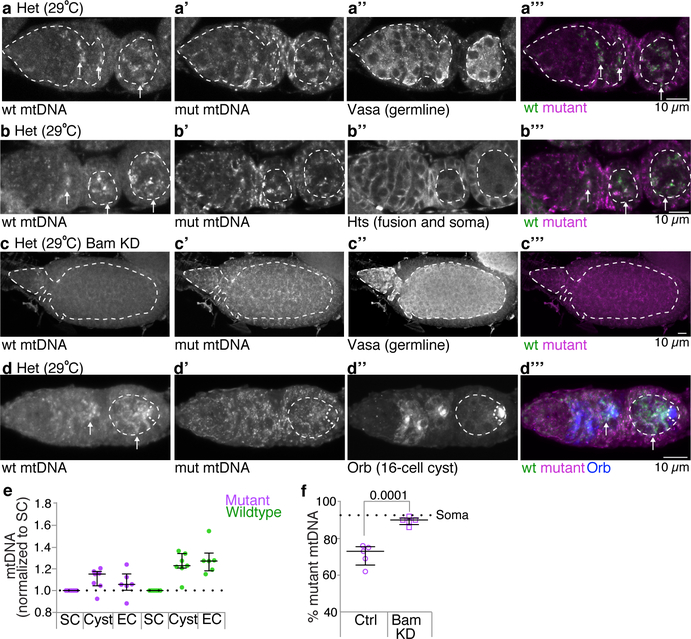
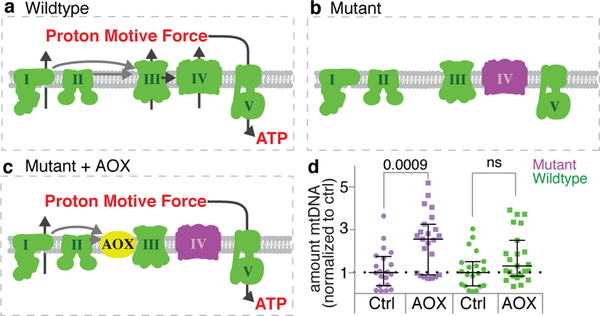
Comment in
-
De-fusing mitochondria defuses the mtDNA time-bomb.Cell Res. 2019 Oct;29(10):781-782. doi: 10.1038/s41422-019-0206-z. Cell Res. 2019. PMID: 31366991 Free PMC article. No abstract available.
-
Genetics: Segregation of Mitochondrial Genomes in the Germline.Curr Biol. 2019 Aug 5;29(15):R746-R748. doi: 10.1016/j.cub.2019.06.029. Curr Biol. 2019. PMID: 31386851
References
Method References
-
- McKim KS, Joyce EF & Jang JK in Methods in Molecular Biology 558, 197–216 (Humana Press, 2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
