

Gaze stabilization in mantis shrimp in response to angled stimuli
- PMID: 31093738
- PMCID: PMC6647723
- DOI: 10.1007/s00359-019-01341-5
Gaze stabilization in mantis shrimp in response to angled stimuli
Abstract
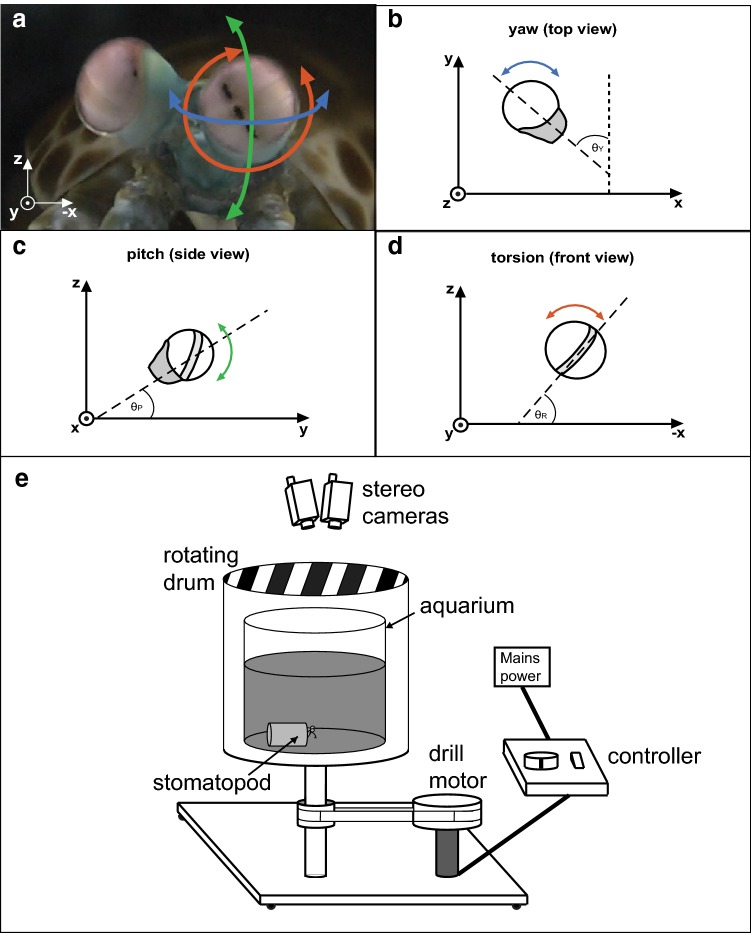
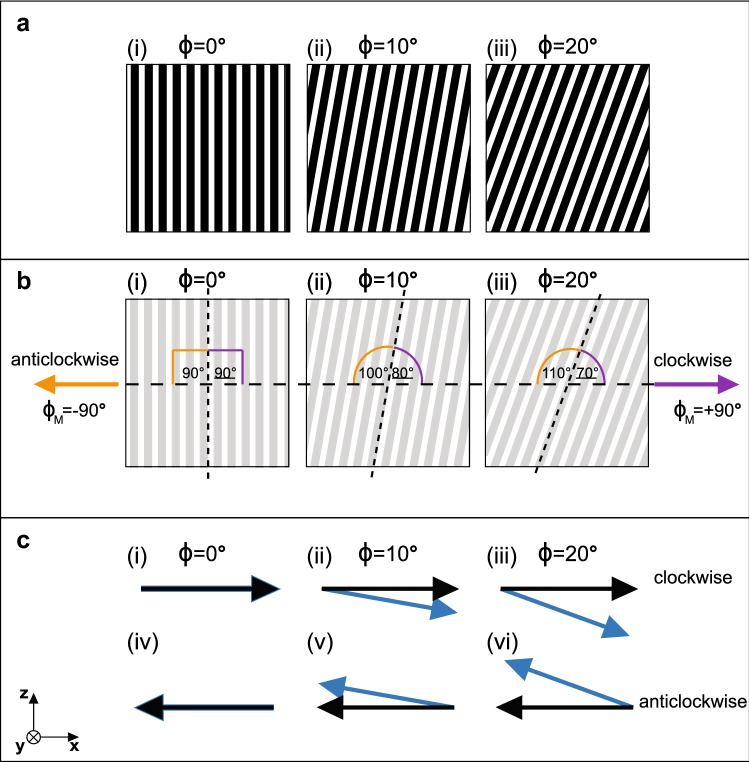
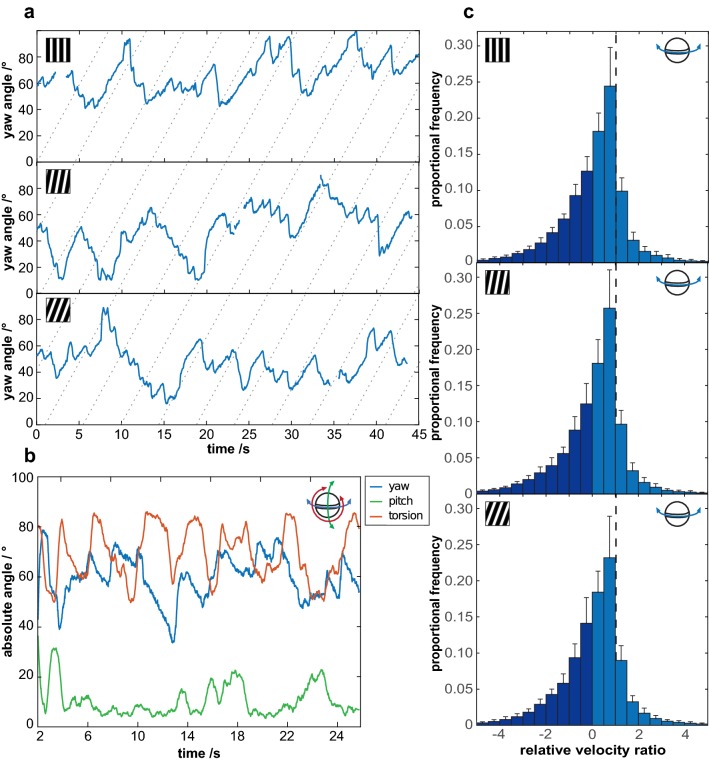
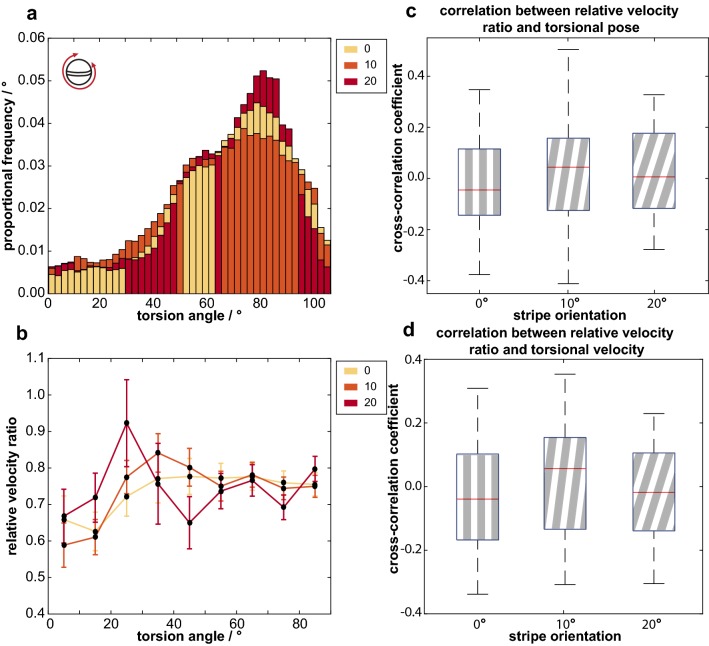
Gaze stabilization is a fundamental aspect of vision and almost all animals shift their eyes to compensate for any self-movement relative to the external environment. When it comes to mantis shrimp, however, the situation becomes complicated due to the complexity of their visual system and their range of eye movements. The stalked eyes of mantis shrimp can independently move left and right, and up and down, whilst simultaneously rotating about the axis of the eye stalks. Despite the large range of rotational freedom, mantis shrimp nevertheless show a stereotypical gaze stabilization response to horizontal motion of a wide-field, high-contrast stimulus. This response is often accompanied by pitch (up-down) and torsion (about the eye stalk) rotations which, surprisingly, have no effect on the performance of yaw (side-to-side) gaze stabilization. This unusual feature of mantis shrimp vision suggests that their neural circuitry for detecting motion is radially symmetric and immune to the confounding effects of torsional self-motion. In this work, we reinforce this finding, demonstrating that the yaw gaze stabilization response of the mantis shrimp is robust to the ambiguous motion cues arising from the motion of striped visual gratings in which the angle of a grating is offset from its direction of travel.
Keywords: Eye movements; Gaze stabilisation; Motion detection; Optokinesis; Stomatopod.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
-
- Bates D, Maechler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015;67:1–48. doi: 10.18637/jss.v067.i01. - DOI
-
- Chatfield C. The analysis of time series: an introduction. 6. Boca Raton: Chapman & Hall/CRC; 2004.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
