

Early Signals of Vaccine-driven Perturbation Seen in Pneumococcal Carriage Population Genomic Data
- PMID: 31094423
- PMCID: PMC7768739
- DOI: 10.1093/cid/ciz404
Early Signals of Vaccine-driven Perturbation Seen in Pneumococcal Carriage Population Genomic Data
Abstract
Background: Pneumococcal conjugate vaccines (PCVs) have reduced pneumococcal diseases globally. Pneumococcal genomic surveys elucidate PCV effects on population structure but are rarely conducted in low-income settings despite the high disease burden.
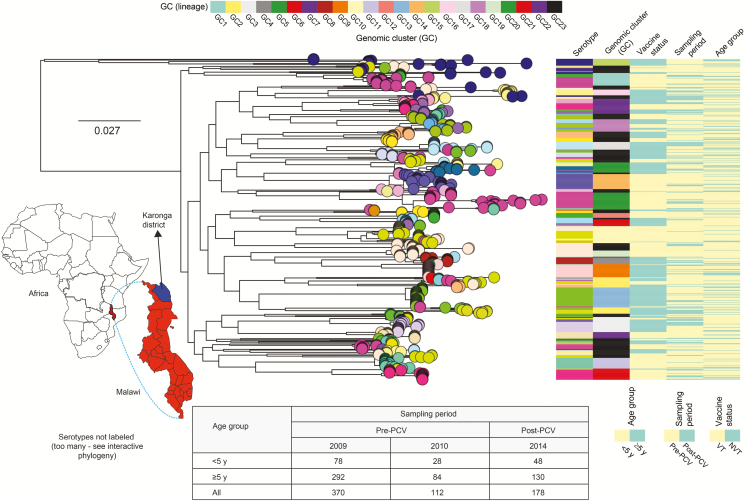
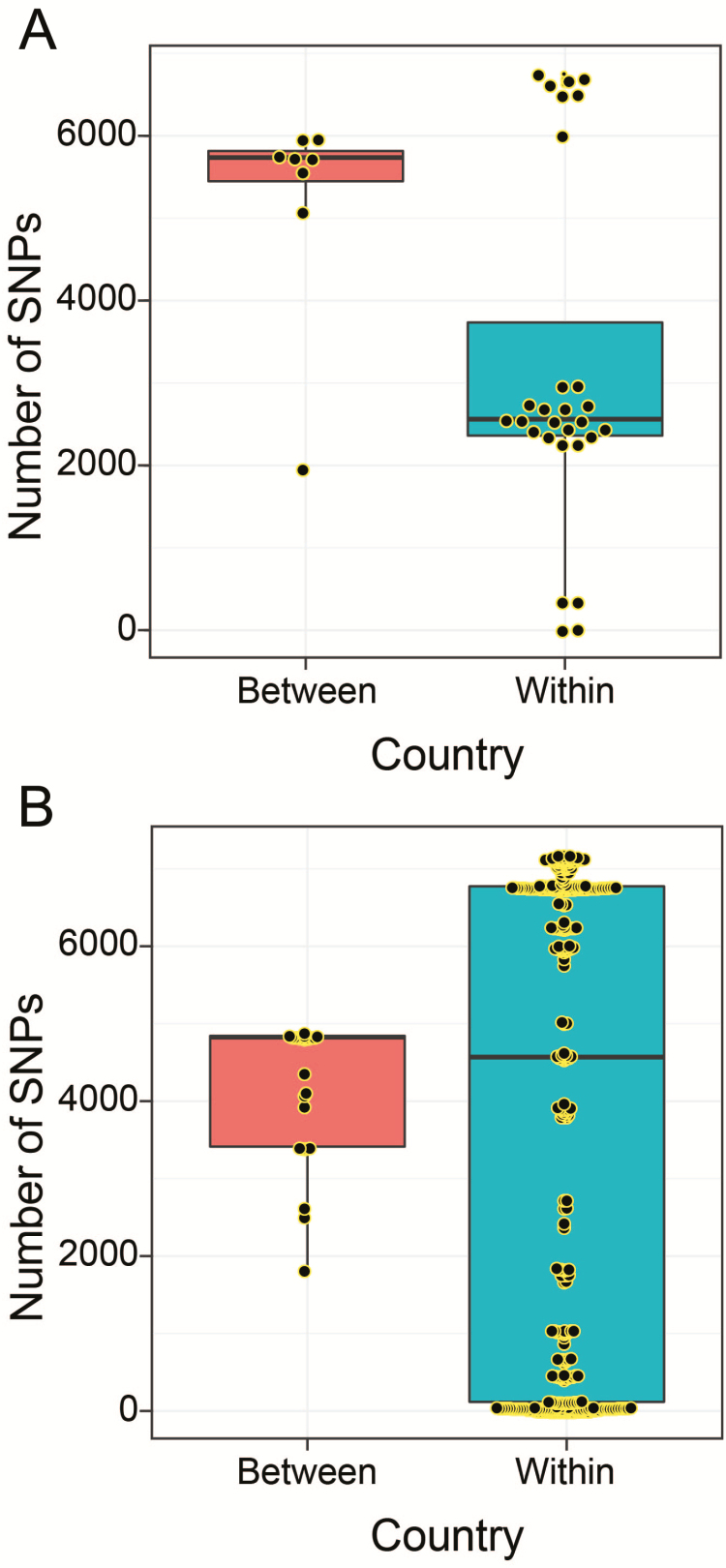
Methods: We undertook whole-genome sequencing (WGS) of 660 pneumococcal isolates collected through surveys from healthy carriers 2 years from 13-valent PCV (PCV13) introduction and 1 year after rollout in northern Malawi. We investigated changes in population structure, within-lineage serotype dynamics, serotype diversity, and frequency of antibiotic resistance (ABR) and accessory genes.
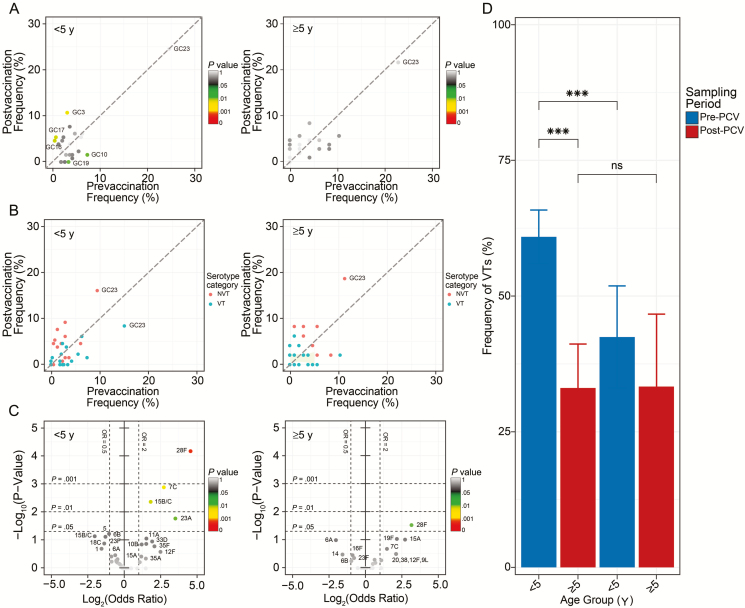
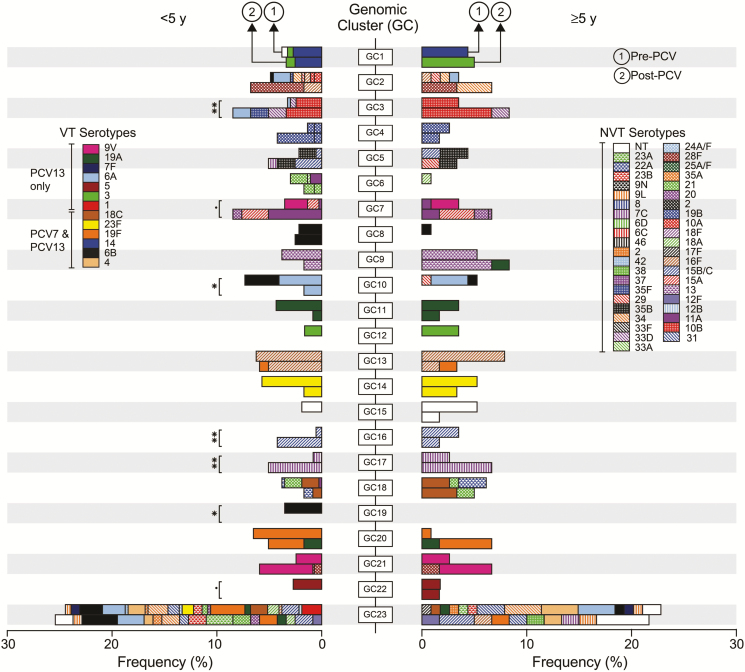
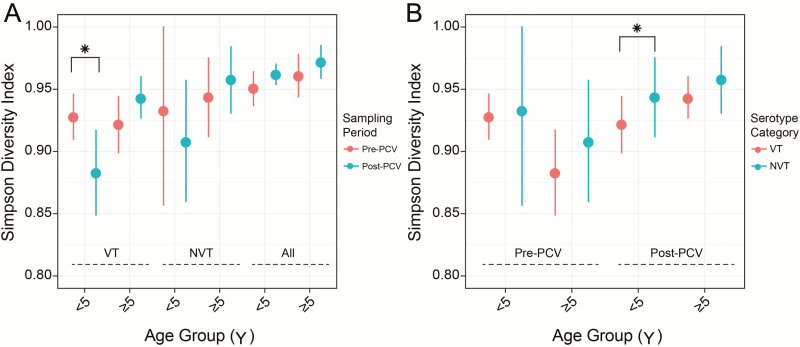
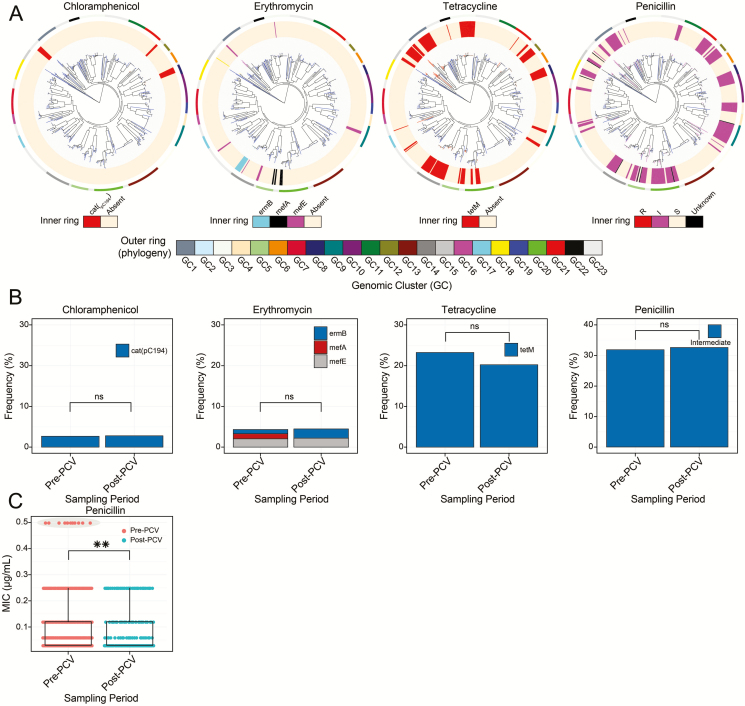
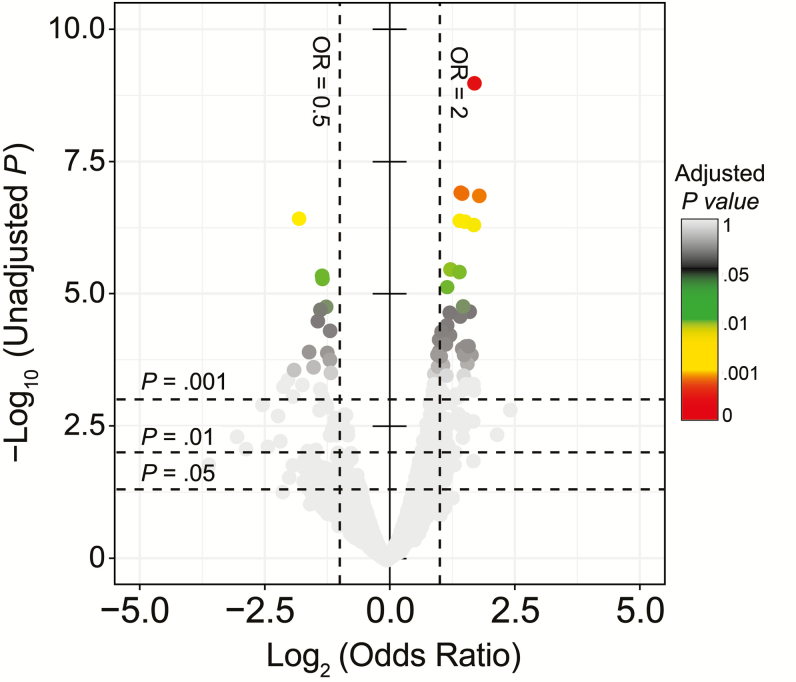
Results: In children <5 years of age, frequency and diversity of vaccine serotypes (VTs) decreased significantly post-PCV, but no significant changes occurred in persons ≥5 years of age. Clearance of VT serotypes was consistent across different genetic backgrounds (lineages). There was an increase of nonvaccine serotypes (NVTs)-namely 7C, 15B/C, and 23A-in children <5 years of age, but 28F increased in both age groups. While carriage rates have been recently shown to remain stable post-PCV due to replacement serotypes, there was no change in diversity of NVTs. Additionally, frequency of intermediate-penicillin-resistant lineages decreased post-PCV. Although frequency of ABR genes remained stable, other accessory genes, especially those associated with mobile genetic element and bacteriocins, showed changes in frequency post-PCV.
Conclusions: We demonstrate evidence of significant population restructuring post-PCV driven by decreasing frequency of vaccine serotypes and increasing frequency of few NVTs mainly in children under 5. Continued surveillance with WGS remains crucial to fully understand dynamics of the residual VTs and replacement NVT serotypes post-PCV.
Keywords: carriage; genomics; pneumococcus; serotypes.
© The Author(s) 2019. Published by Oxford University Press for the Infectious Diseases Society of America.
Figures
References
-
- Ben-Shimol S, Givon-Lavi N, Leibovitz E, Raiz S, Greenberg D, Dagan R. Impact of widespread introduction of pneumococcal conjugate vaccines on pneumococcal and nonpneumococcal otitis media. Clin Infect Dis 2016; 63:611–8. - PubMed
-
- Whitney CG, Farley MM, Hadler J, et al. Active Bacterial Core Surveillance of the Emerging Infections Program Network Decline in invasive pneumococcal disease after the introduction of protein-polysaccharide conjugate vaccine. N Engl J Med 2003; 348:1737–46. - PubMed
-
- von Gottberg A, de Gouveia L, Tempia S, et al. GERMS-SA Investigators Effects of vaccination on invasive pneumococcal disease in South Africa. N Engl J Med 2014; 371:1889–99. - PubMed
-
- Hammitt LL, Akech DO, Morpeth SC, et al. Population effect of 10-valent pneumococcal conjugate vaccine on nasopharyngeal carriage of Streptococcus pneumoniae and non-typeable Haemophilus influenzae in Kilifi, Kenya: findings from cross-sectional carriage studies. Lancet Glob Health 2014; 2:e397–405. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
