

Reading canonical and modified nucleobases in 16S ribosomal RNA using nanopore native RNA sequencing
- PMID: 31095620
- PMCID: PMC6522004
- DOI: 10.1371/journal.pone.0216709
Reading canonical and modified nucleobases in 16S ribosomal RNA using nanopore native RNA sequencing
Abstract
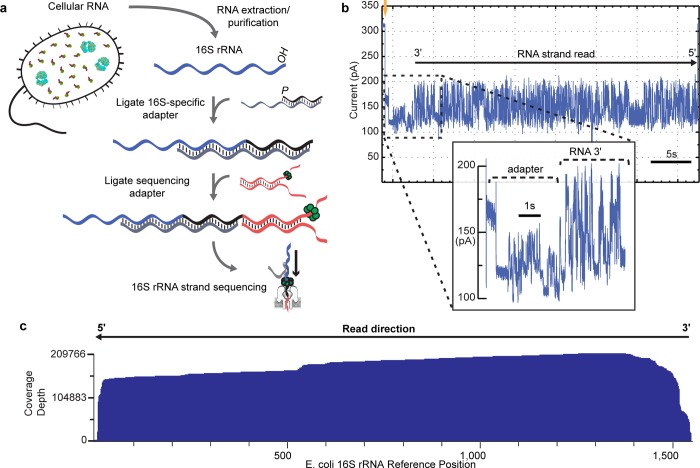
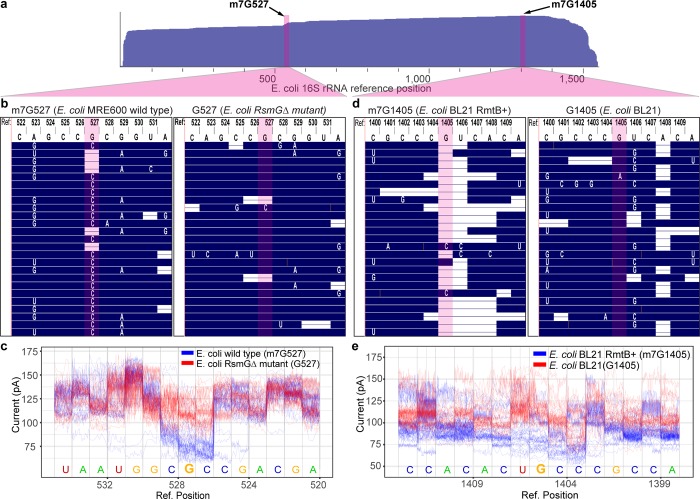
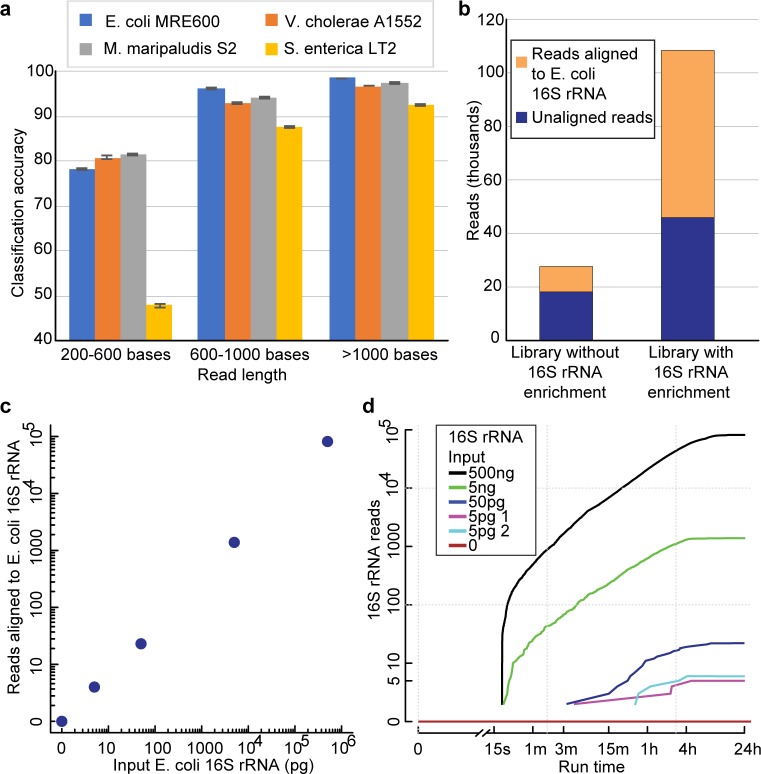
The ribosome small subunit is expressed in all living cells. It performs numerous essential functions during translation, including formation of the initiation complex and proofreading of base-pairs between mRNA codons and tRNA anticodons. The core constituent of the small ribosomal subunit is a ~1.5 kb RNA strand in prokaryotes (16S rRNA) and a homologous ~1.8 kb RNA strand in eukaryotes (18S rRNA). Traditional sequencing-by-synthesis (SBS) of rRNA genes or rRNA cDNA copies has achieved wide use as a 'molecular chronometer' for phylogenetic studies, and as a tool for identifying infectious organisms in the clinic. However, epigenetic modifications on rRNA are erased by SBS methods. Here we describe direct MinION nanopore sequencing of individual, full-length 16S rRNA absent reverse transcription or amplification. As little as 5 picograms (~10 attomole) of purified E. coli 16S rRNA was detected in 4.5 micrograms of total human RNA. Nanopore ionic current traces that deviated from canonical patterns revealed conserved E. coli 16S rRNA 7-methylguanosine and pseudouridine modifications, and a 7-methylguanosine modification that confers aminoglycoside resistance to some pathological E. coli strains.
Conflict of interest statement
MA holds options in Oxford Nanopore Technologies (ONT). MA is a paid consultant to ONT. MA is an inventor on 11 University of California patents licensed to ONT (6,267,872, 6,465,193, 6,746,594, 6,936,433, 7,060,50, 8,500,982, 8,679,747, 9,481,908, 9,797,013, 10,059,988, and 10,081,835). DRG, who contributed to each facet of the paper, is an employee of Oxford Nanopore Technologies. This does not alter our adherence to PLOS ONE policies on sharing data and materials.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
