

TPX2-LIKE PROTEIN3 Is the Primary Activator of α-Aurora Kinases and Is Essential for Embryogenesis
- PMID: 31097675
- PMCID: PMC6752915
- DOI: 10.1104/pp.18.01515
TPX2-LIKE PROTEIN3 Is the Primary Activator of α-Aurora Kinases and Is Essential for Embryogenesis
Abstract
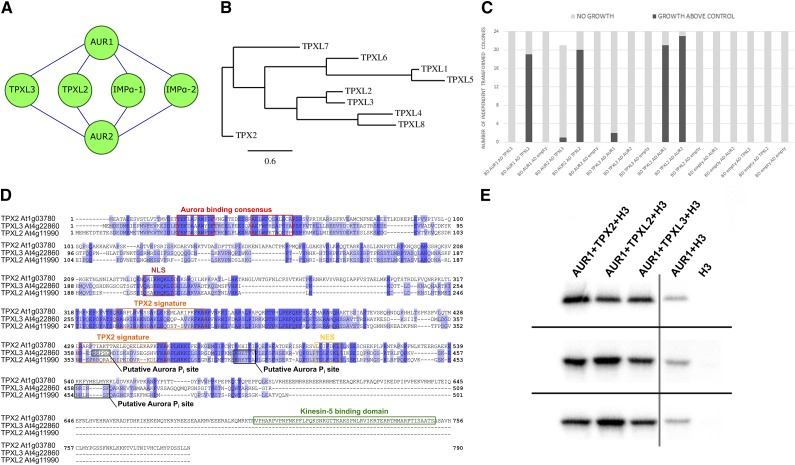
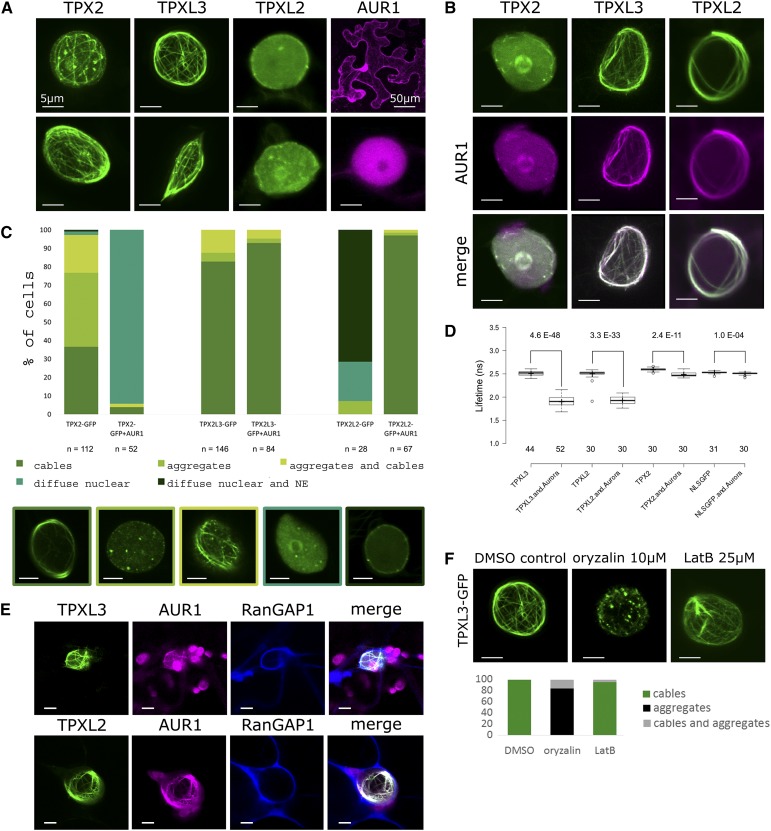
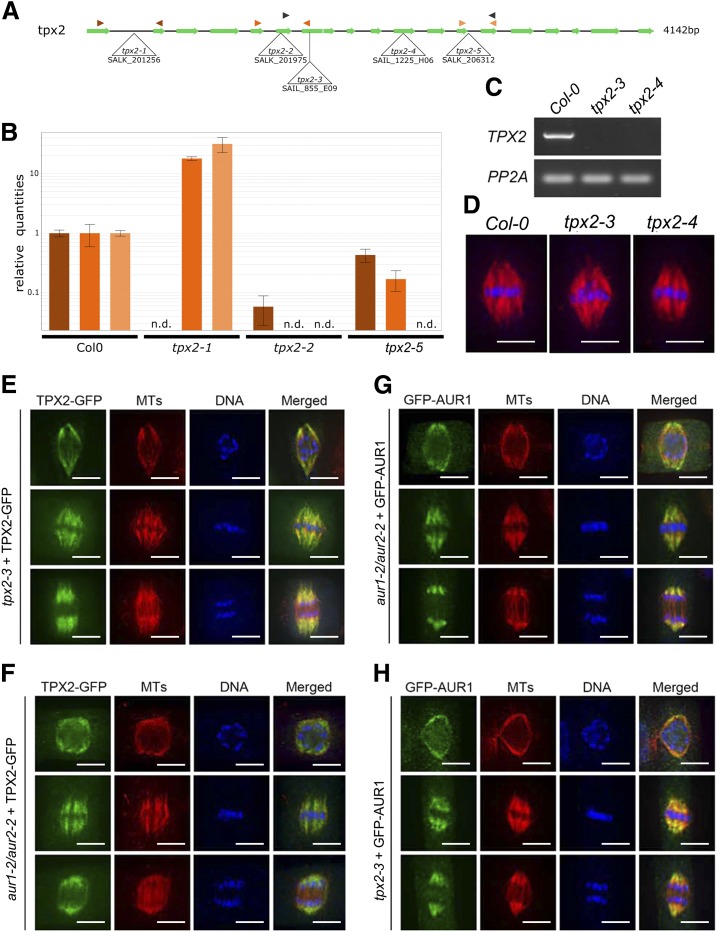
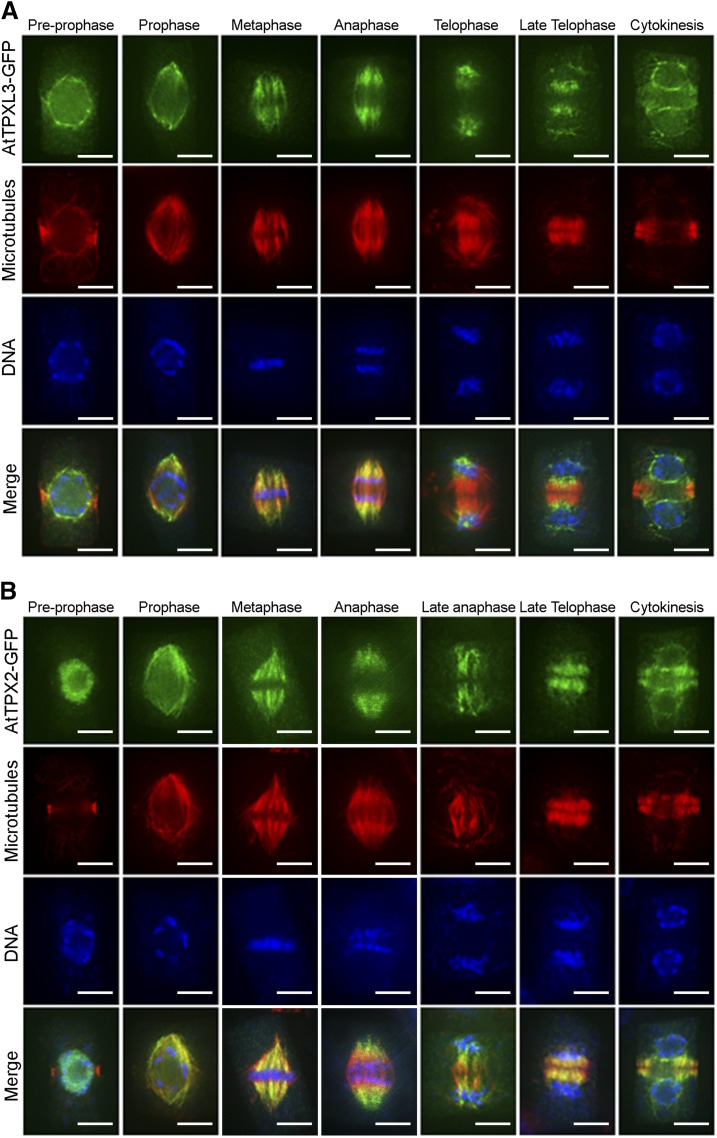
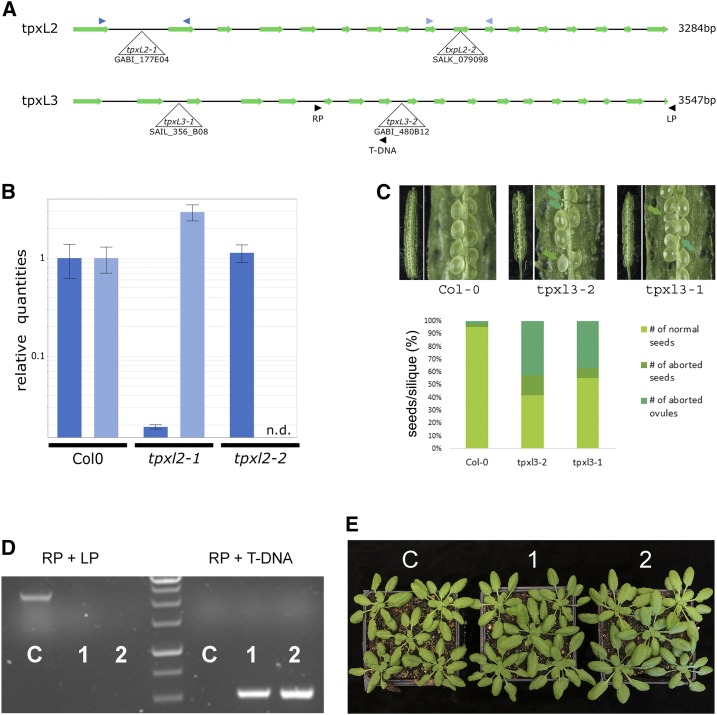
Aurora kinases are key regulators of mitosis. Multicellular eukaryotes generally possess two functionally diverged types of Aurora kinases. In plants, including Arabidopsis (Arabidopsis thaliana), these are termed α- and β-Auroras. As the functional specification of Aurora kinases is determined by their specific interaction partners, we initiated interactomics analyses using both Arabidopsis α-Aurora kinases (AUR1 and AUR2). Proteomics results revealed that TPX2-LIKE PROTEINS2 and 3 (TPXL2/3) prominently associated with α-Auroras, as did the conserved TPX2 to a lower degree. Like TPX2, TPXL2 and TPXL3 strongly activated the AUR1 kinase but exhibited cell-cycle-dependent localization differences on microtubule arrays. The separate functions of TPX2 and TPXL2/3 were also suggested by their different influences on AUR1 localization upon ectopic expressions. Furthermore, genetic analyses showed that TPXL3, but not TPX2 and TPXL2, acts nonredundantly to enable proper embryo development. In contrast to vertebrates, plants have an expanded TPX2 family and these family members have both redundant and unique functions. Moreover, as neither TPXL2 nor TPXL3 contains the C-terminal Kinesin-5 binding domain present in the canonical TPX2, the targeting and activity of this kinesin must be organized differently in plants.
© 2019 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Barnett J. (1991) Microtubules in interphase nuclei of Aesculus hippocastanum L. Ann Bot (Lond) 68: 159–165
-
- Barr AR, Gergely F (2007) Aurora-A: The maker and breaker of spindle poles. J Cell Sci 120: 2987–2996 - PubMed
-
- Bayliss R, Sardon T, Vernos I, Conti E (2003) Structural basis of Aurora-A activation by TPX2 at the mitotic spindle. Mol Cell 12: 851–862 - PubMed
-
- Bayliss R, Sardon T, Ebert J, Lindner D, Vernos I, Conti E (2004) Determinants for Aurora-A activation and Aurora-B discrimination by TPX2. Cell Cycle 3: 404–407 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
