

Yin Yang 1 sustains biosynthetic demands during brain development in a stage-specific manner
- PMID: 31097699
- PMCID: PMC6522535
- DOI: 10.1038/s41467-019-09823-5
Yin Yang 1 sustains biosynthetic demands during brain development in a stage-specific manner
Abstract
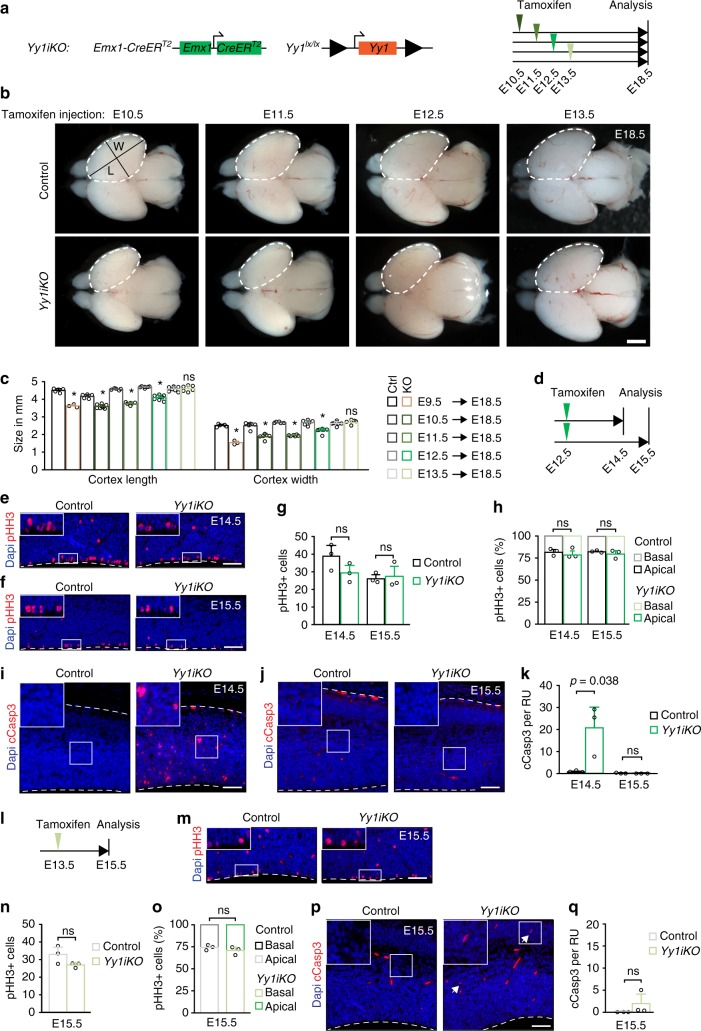
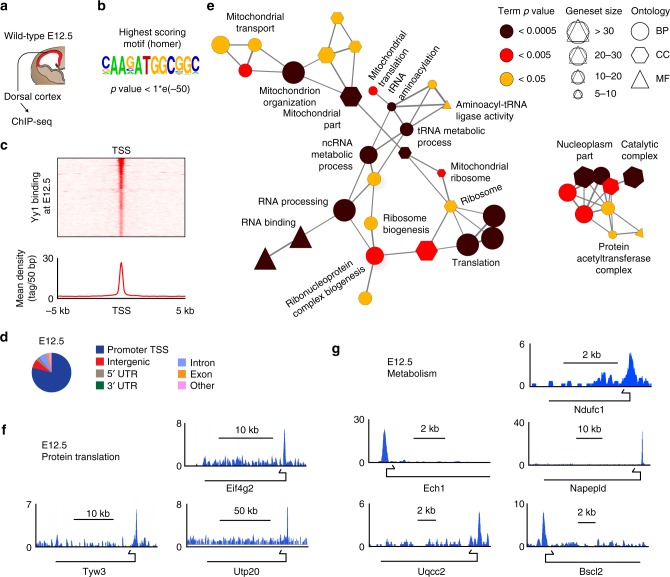
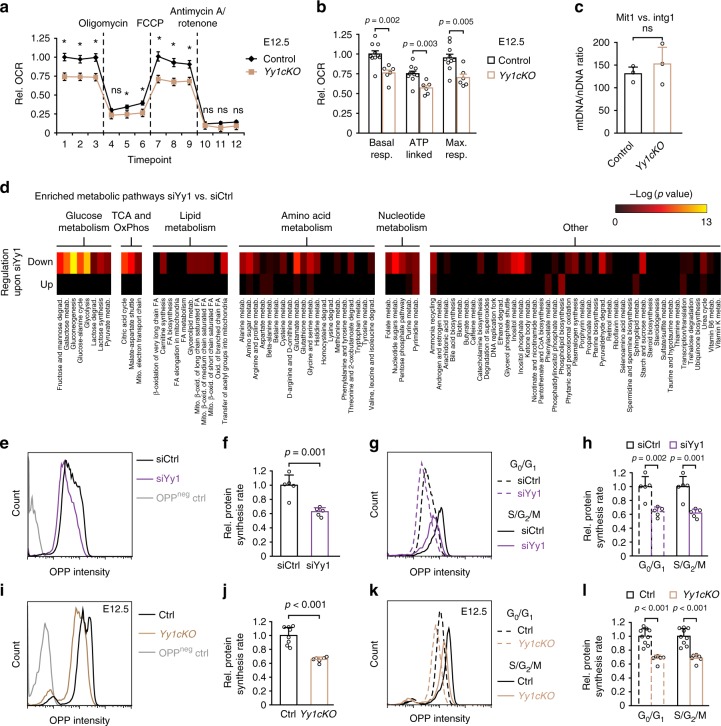
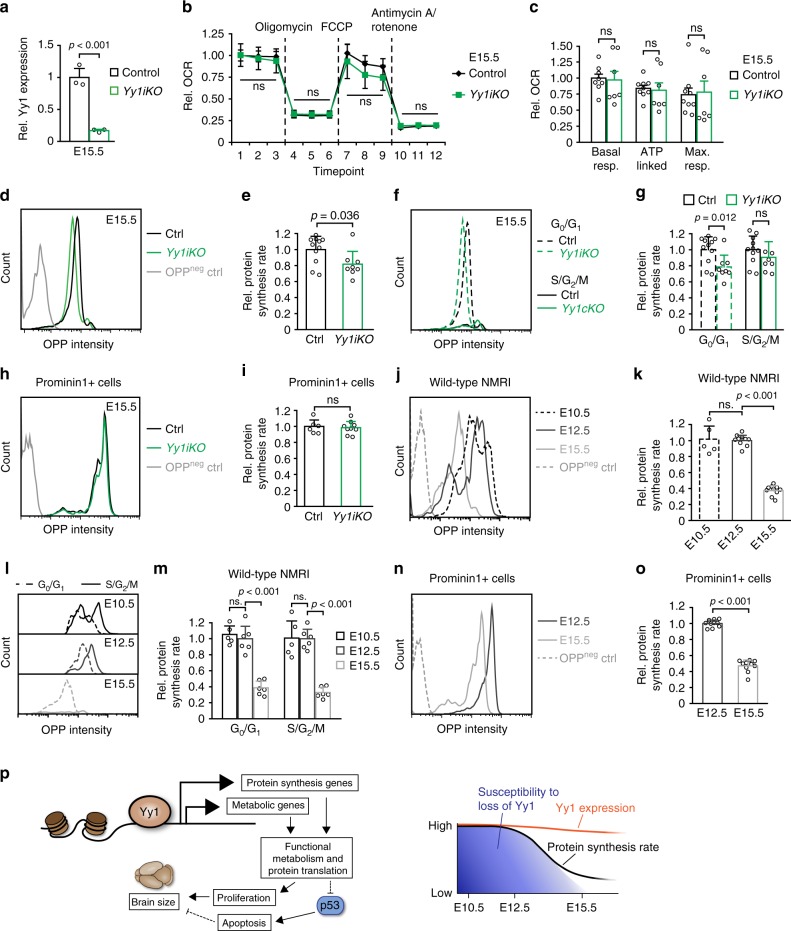
The transcription factor Yin Yang 1 (YY1) plays an important role in human disease. It is often overexpressed in cancers and mutations can lead to a congenital haploinsufficiency syndrome characterized by craniofacial dysmorphisms and neurological dysfunctions, consistent with a role in brain development. Here, we show that Yy1 controls murine cerebral cortex development in a stage-dependent manner. By regulating a wide range of metabolic pathways and protein translation, Yy1 maintains proliferation and survival of neural progenitor cells (NPCs) at early stages of brain development. Despite its constitutive expression, however, the dependence on Yy1 declines over the course of corticogenesis. This is associated with decreasing importance of processes controlled by Yy1 during development, as reflected by diminished protein synthesis rates at later developmental stages. Thus, our study unravels a novel role for Yy1 as a stage-dependent regulator of brain development and shows that biosynthetic demands of NPCs dynamically change throughout development.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
