

Antiviral RNAi in Insects and Mammals: Parallels and Differences
- PMID: 31100912
- PMCID: PMC6563508
- DOI: 10.3390/v11050448
Antiviral RNAi in Insects and Mammals: Parallels and Differences
Abstract
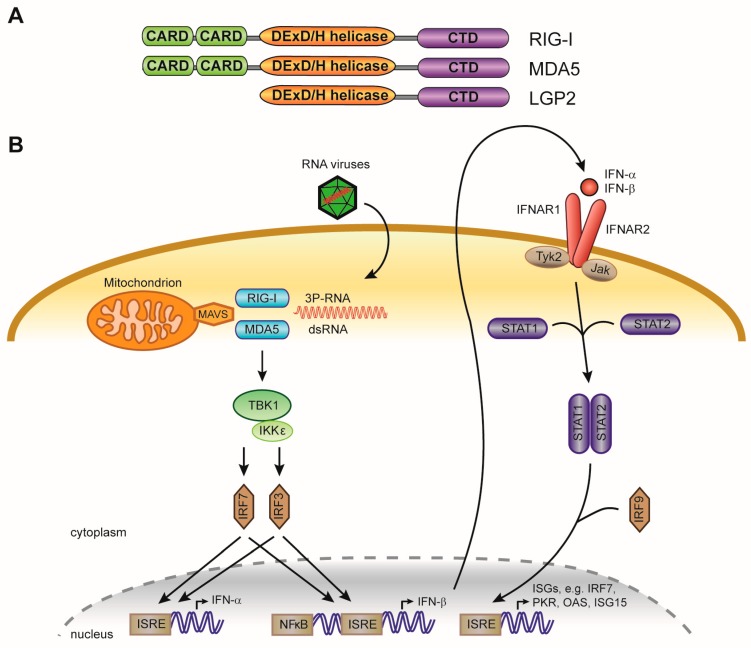
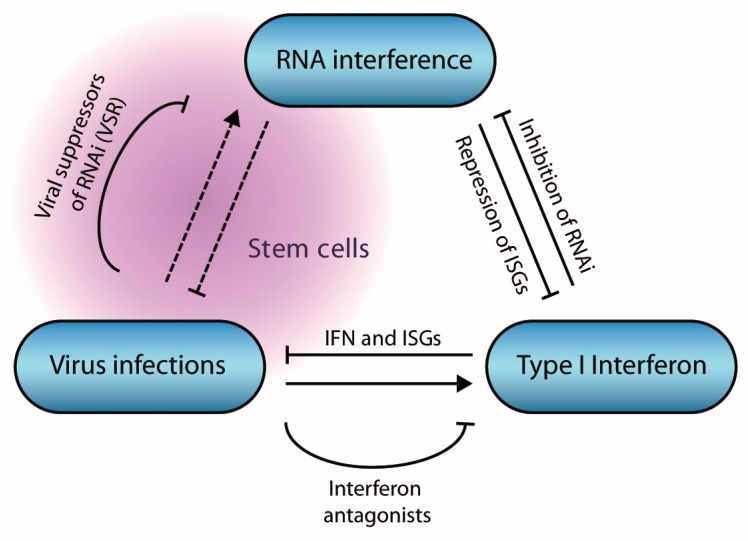
The RNA interference (RNAi) pathway is a potent antiviral defense mechanism in plants and invertebrates, in response to which viruses evolved suppressors of RNAi. In mammals, the first line of defense is mediated by the type I interferon system (IFN); however, the degree to which RNAi contributes to antiviral defense is still not completely understood. Recent work suggests that antiviral RNAi is active in undifferentiated stem cells and that antiviral RNAi can be uncovered in differentiated cells in which the IFN system is inactive or in infections with viruses lacking putative viral suppressors of RNAi. In this review, we describe the mechanism of RNAi and its antiviral functions in insects and mammals. We draw parallels and highlight differences between (antiviral) RNAi in these classes of animals and discuss open questions for future research.
Keywords: RNA interference; RNA virus; antiviral defense; innate immunity; interferon; small interfering RNA; stem cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
RNAi-mediated antiviral immunity in mammals.Curr Opin Virol. 2018 Oct;32:9-14. doi: 10.1016/j.coviro.2018.07.008. Epub 2018 Jul 14. Curr Opin Virol. 2018. PMID: 30015014 Review.
-
RNAi-mediated antiviral immunity in insects and their possible application.Curr Opin Virol. 2018 Oct;32:108-114. doi: 10.1016/j.coviro.2018.10.004. Epub 2018 Nov 12. Curr Opin Virol. 2018. PMID: 30428425 Review.
-
Antiviral innate immune response of RNA interference.J Infect Dev Ctries. 2014 Jul 14;8(7):804-10. doi: 10.3855/jidc.4187. J Infect Dev Ctries. 2014. PMID: 25022288 Review.
-
The Interplay Between Viruses and RNAi Pathways in Insects.Annu Rev Entomol. 2021 Jan 7;66:61-79. doi: 10.1146/annurev-ento-033020-090410. Annu Rev Entomol. 2021. PMID: 33417818 Review.
-
Bugs Are Not to Be Silenced: Small RNA Pathways and Antiviral Responses in Insects.Annu Rev Virol. 2016 Sep 29;3(1):573-589. doi: 10.1146/annurev-virology-110615-042447. Annu Rev Virol. 2016. PMID: 27741406 Review.
Cited by
-
Host Factors That Control Mosquito-Borne Viral Infections in Humans and Their Vector.Viruses. 2021 Apr 24;13(5):748. doi: 10.3390/v13050748. Viruses. 2021. PMID: 33923307 Free PMC article. Review.
-
The Aedes aegypti siRNA pathway mediates broad-spectrum defense against human pathogenic viruses and modulates antibacterial and antifungal defenses.PLoS Biol. 2022 Jun 9;20(6):e3001668. doi: 10.1371/journal.pbio.3001668. eCollection 2022 Jun. PLoS Biol. 2022. PMID: 35679279 Free PMC article.
-
An RNA Interference/Adeno-Associated Virus Vector-Based Combinatorial Gene Therapy Approach Against Hepatitis E Virus.Hepatol Commun. 2022 Apr;6(4):878-888. doi: 10.1002/hep4.1842. Epub 2021 Oct 31. Hepatol Commun. 2022. PMID: 34719133 Free PMC article.
-
Endogenous Feline Leukemia Virus (FeLV) siRNA Transcription May Interfere with Exogenous FeLV Infection.J Virol. 2021 Nov 9;95(23):e0007021. doi: 10.1128/JVI.00070-21. Epub 2021 Sep 8. J Virol. 2021. PMID: 34495702 Free PMC article.
-
Oropouche orthobunyavirus in Urban Mosquitoes: Vector Competence, Coinfection, and Immune System Activation in Aedes aegypti.Viruses. 2025 Mar 28;17(4):492. doi: 10.3390/v17040492. Viruses. 2025. PMID: 40284935 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
