

Posttranscription Initiation Control of Gene Expression Mediated by Bacterial RNA-Binding Proteins
- PMID: 31100987
- PMCID: PMC9404307
- DOI: 10.1146/annurev-micro-020518-115907
Posttranscription Initiation Control of Gene Expression Mediated by Bacterial RNA-Binding Proteins
Abstract
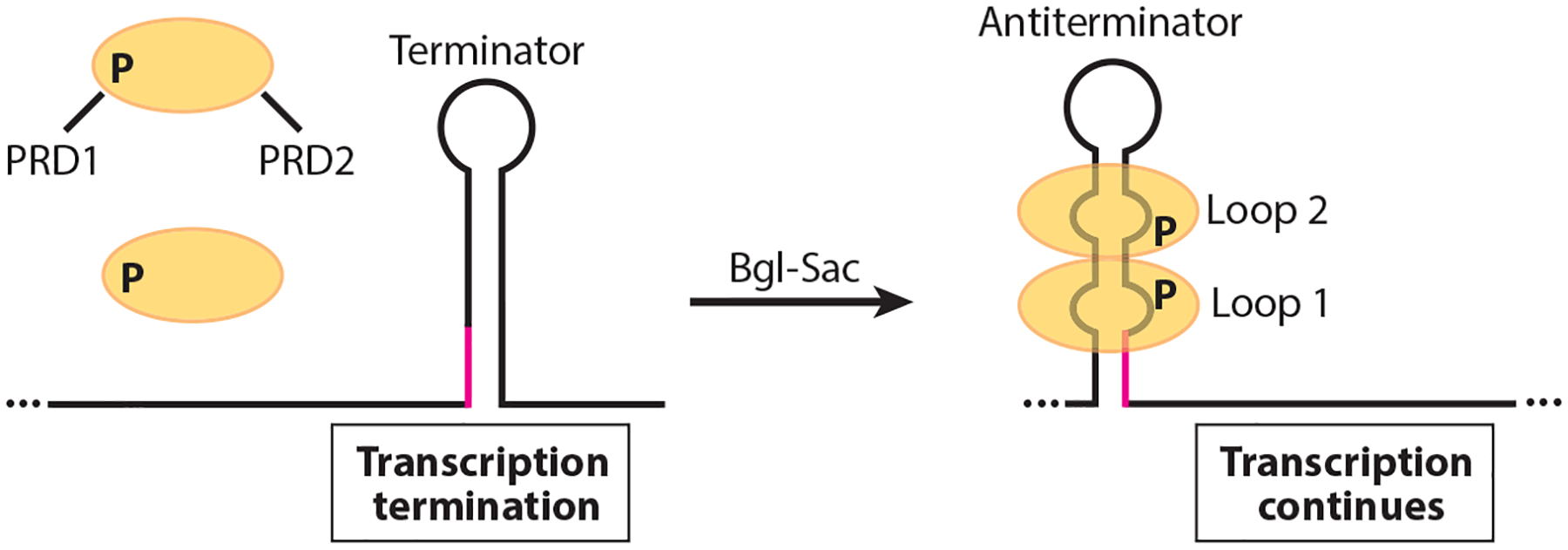
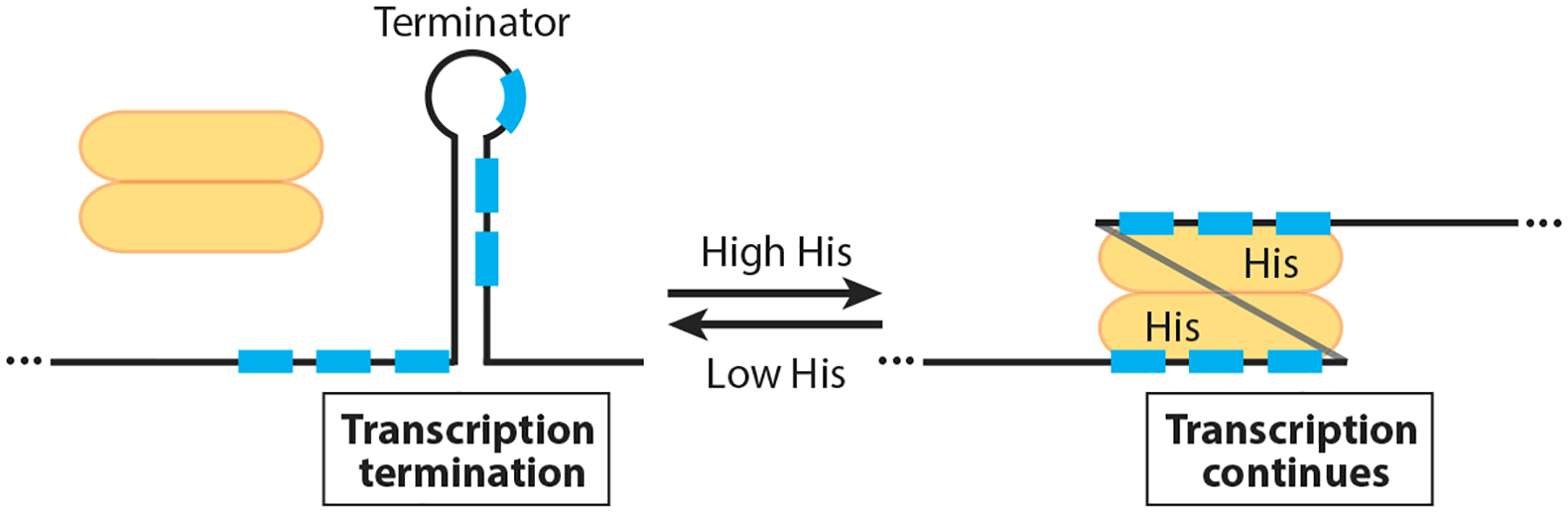
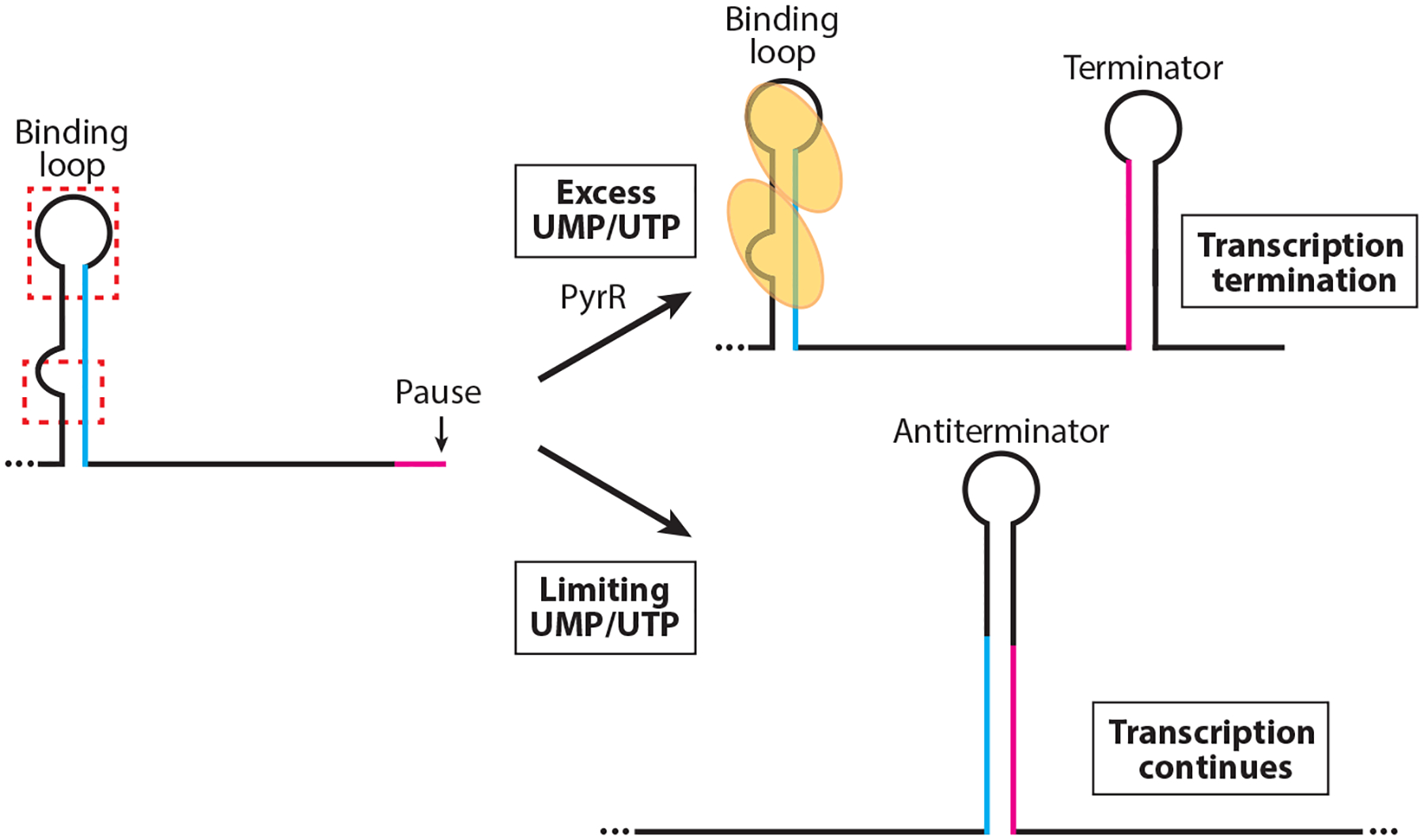
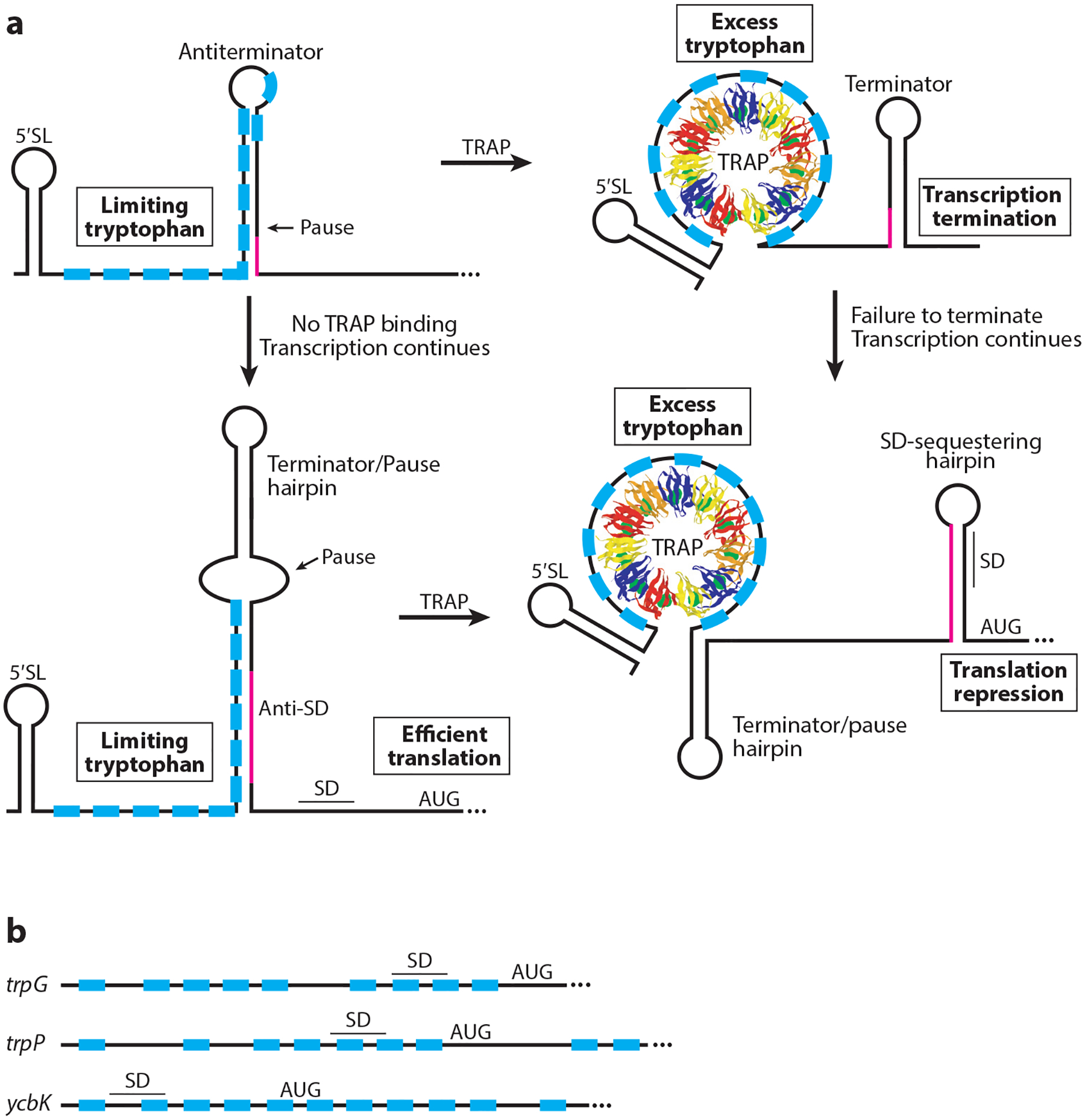
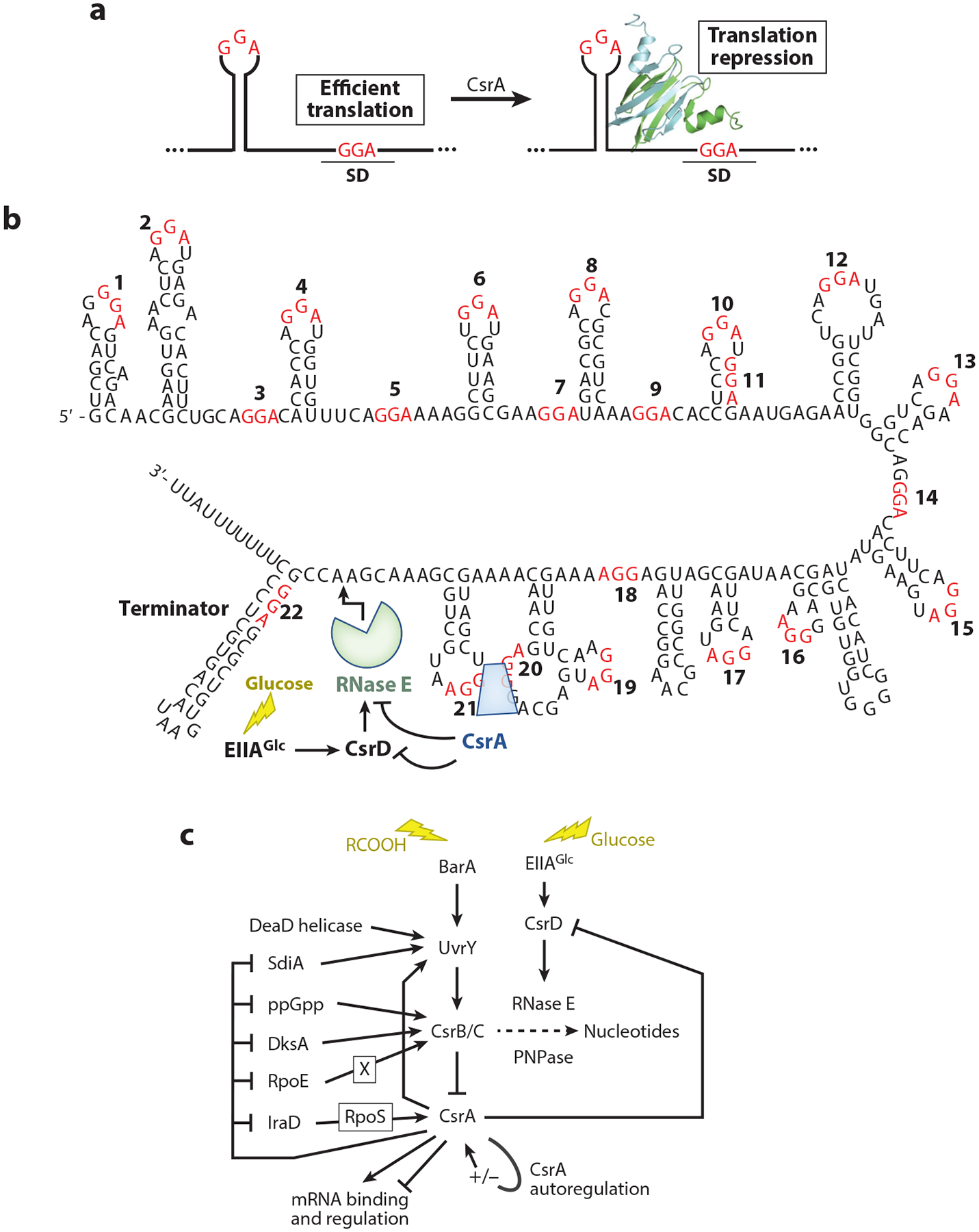
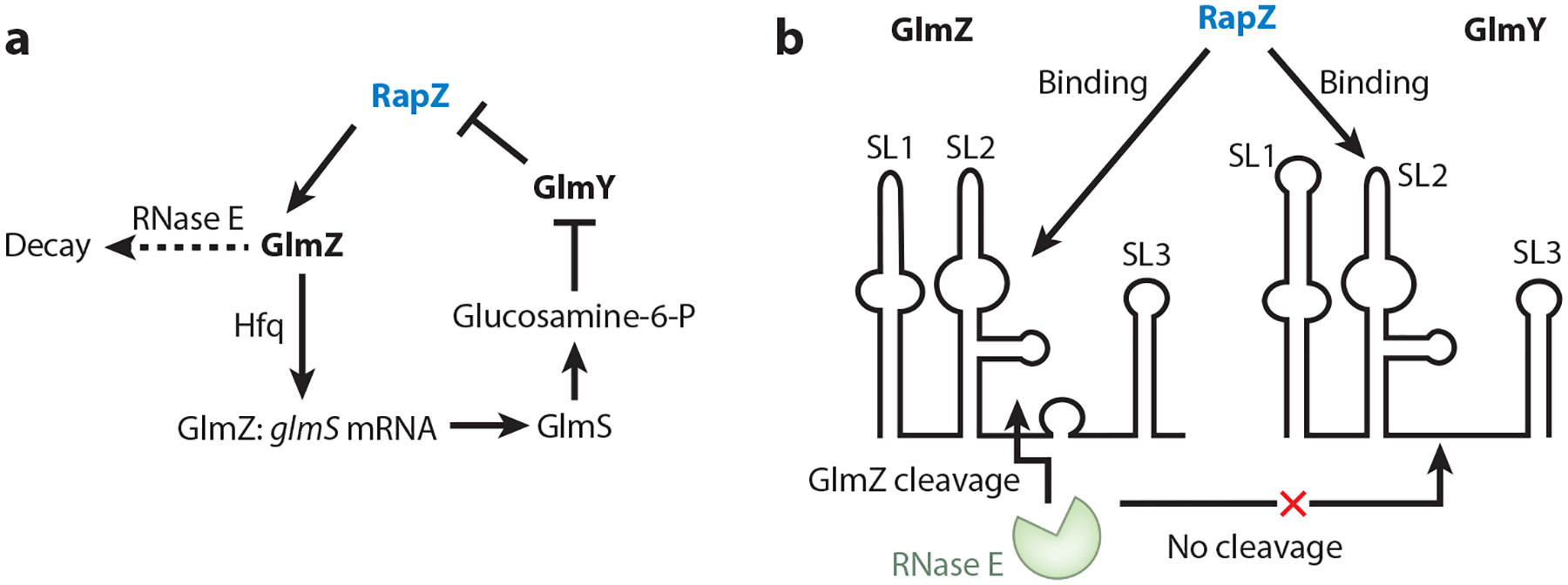
RNA-binding proteins play vital roles in regulating gene expression and cellular physiology in all organisms. Bacterial RNA-binding proteins can regulate transcription termination via attenuation or antitermination mechanisms, while others can repress or activate translation initiation by affecting ribosome binding. The RNA targets for these proteins include short repeated sequences, longer single-stranded sequences, RNA secondary or tertiary structure, and a combination of these features. The activity of these proteins can be influenced by binding of metabolites, small RNAs, or other proteins, as well as by phosphorylation events. Some of these proteins regulate specific genes, while others function as global regulators. As the regulatory mechanisms, components, targets, and signaling circuitry surrounding RNA-binding proteins have become better understood, in part through rapid advances provided by systems approaches, a sense of the true nature of biological complexity is becoming apparent, which we attempt to capture for the reader of this review.
Keywords: RNA-binding protein; antitermination; attenuation; gene regulation; sRNA; translation.
Figures

References
-
- Amster-Choder O 2005. The bgl sensory system: a transmembrane signaling pathway controlling transcriptional antitermination. Curr. Opin. Microbiol 8:127–34 - PubMed
-
- Amster-Choder O, Houman F, Wright A. 1989. Protein phosphorylation regulates transcription of the β-glucoside utilization operon in E. coli. Cell 58:847–55 - PubMed
-
- Amster-Choder O, Wright A. 1992. Modulation of the dimerization of a transcriptional antiterminator protein by phosphorylation. Science 257:1395–98 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
