

Quantitative organization of the excitatory synapses of the primate cerebellar nuclei: further evidence for a specialized architecture underlying the primate cerebellum
- PMID: 31101974
- PMCID: PMC6591209
- DOI: 10.1007/s00429-019-01888-8
Quantitative organization of the excitatory synapses of the primate cerebellar nuclei: further evidence for a specialized architecture underlying the primate cerebellum
Abstract
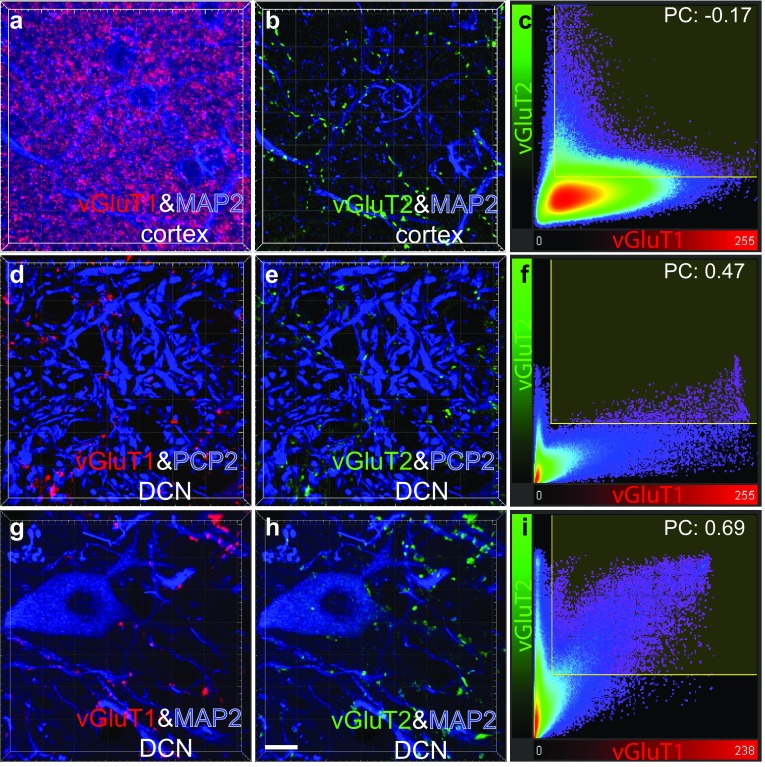
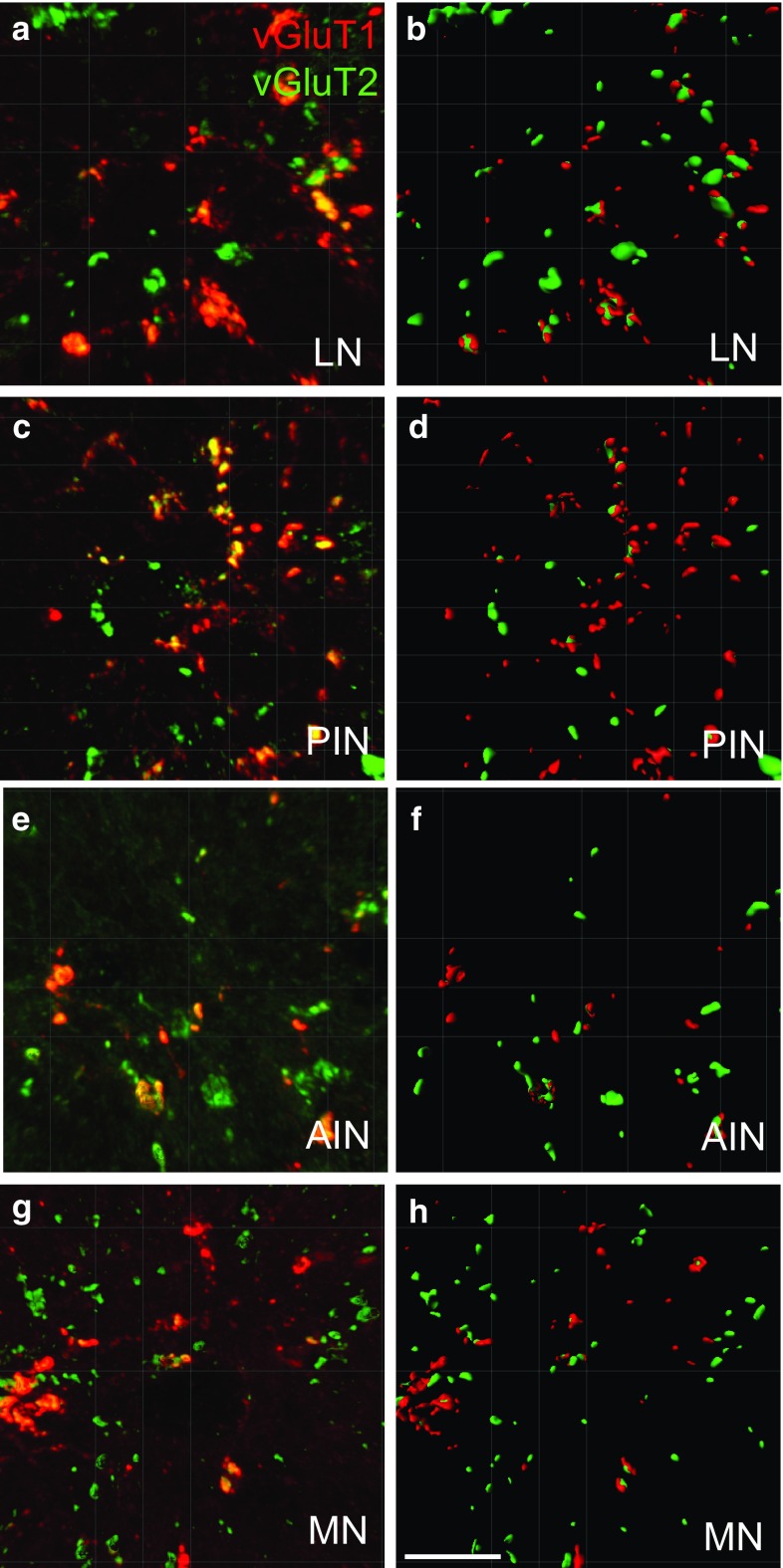
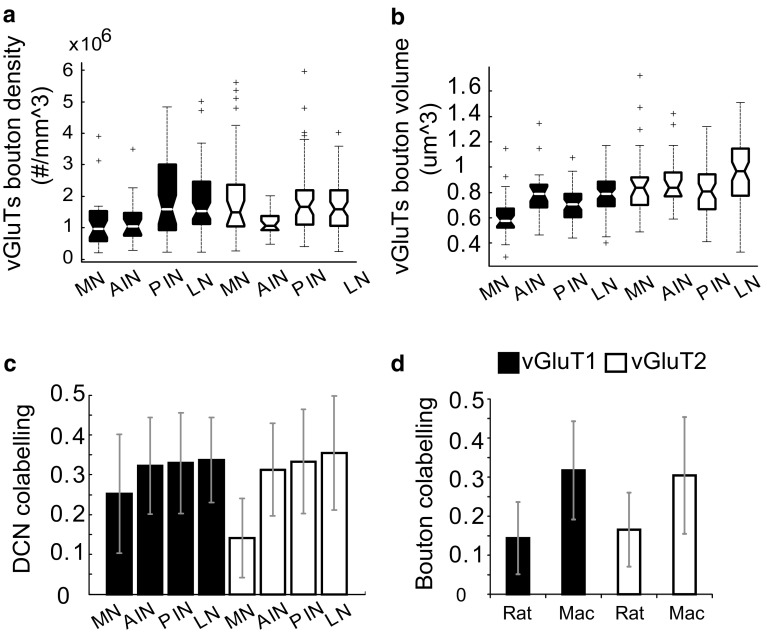
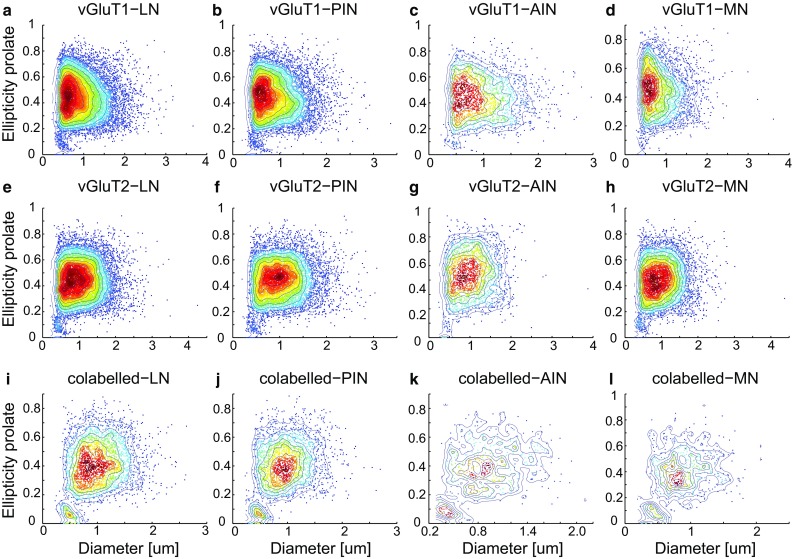
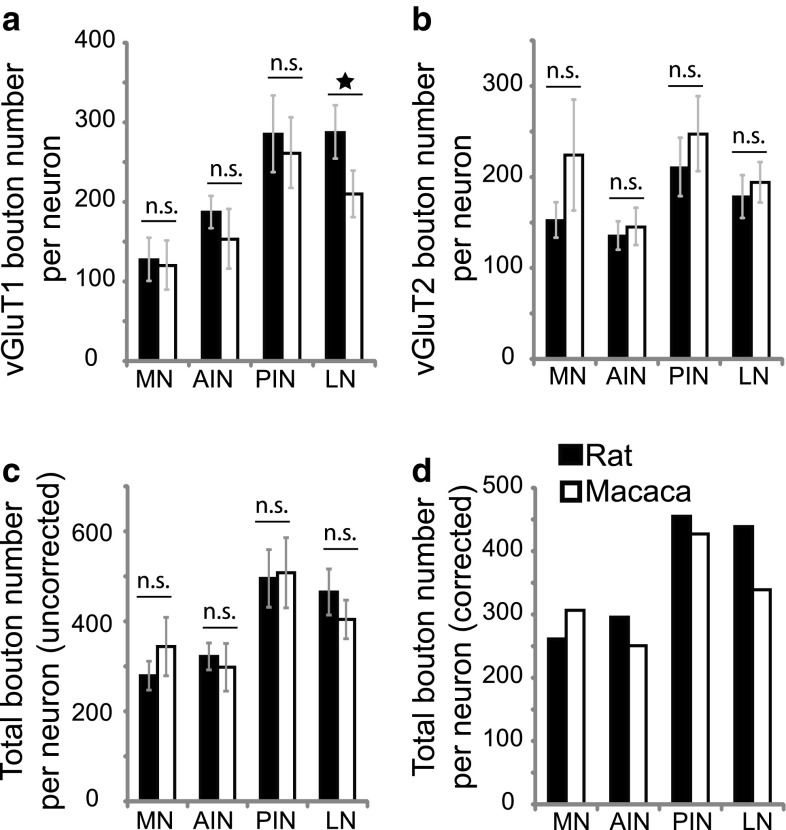
The cerebellar intrinsic connectivity is of remarkable regularity with a similar build repeated many times over. However, several modifications of this basic circuitry occur that can provide important clues to evolutionary adaptations. We have observed differences in the wiring of the cerebellar output structures (the deep cerebellar nuclei, DCN) with higher dendritic length density in the phylogenetically newer DCN. In rats, we showed that an increase in wiring is associated with an increase in the presynaptic vesicular glutamate transporter 1 (vGluT1). In this study, we have extended our analysis to the rhesus monkey and can show similarities and differences between the two species. The similarities confirm a higher density in vGluT1+ boutons in the lateral (LN/dentate) and posterior interpositus nucleus compared to the phylogenetically older DCN. In general, we also observe a lower density of vGluT1 and 2+ boutons in the monkey, which however, yields a similar number of excitatory boutons per neuron in both species. The only exception is the vGlut1+ boutons in the macaque LN/dentate, which showed a significantly lower number of vGluT1+ boutons per neuron. We also detected a higher percentage of co-labelled vGluT1 and 2 boutons in the macaque than we found in the rat. In summary, these results confirm that the hyposcalled dendrites of the monkey LN/dentate also show a lower number of vGluT1+ boutons per neuron. These results provide further support of our model relating the dendritic morphology of the LN/dentate neurons to the morphology of the specially enlarged LN/dentate nucleus in primates.
Keywords: Comparative neuroanatomy; Deep cerebellar nuclei; Lipofuscin fluorescence removal; Quantitative immunofluorescence; Vesicular glutamate transporter.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Boulland JL, Qureshi T, Seal RP, Rafiki A, Gundersen V, Bergersen LH, Fremeau RT, Jr, Edwards RH, Storm-Mathisen J, Chaudhry FA. Expression of the vesicular glutamate transporters during development indicates the widespread corelease of multiple neurotransmitters. J Comp Neurol. 2004;480:264–280. doi: 10.1002/cne.20354. - DOI - PubMed
-
- Braitenberg V, Schüz A. Cortex: statistics and geometry of neuronal connectivity. Berlin: Springer; 1998.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
