

Function-related conformational dynamics of G protein-coupled receptors revealed by NMR
- PMID: 31102199
- PMCID: PMC6557943
- DOI: 10.1007/s12551-019-00539-w
Function-related conformational dynamics of G protein-coupled receptors revealed by NMR
Abstract
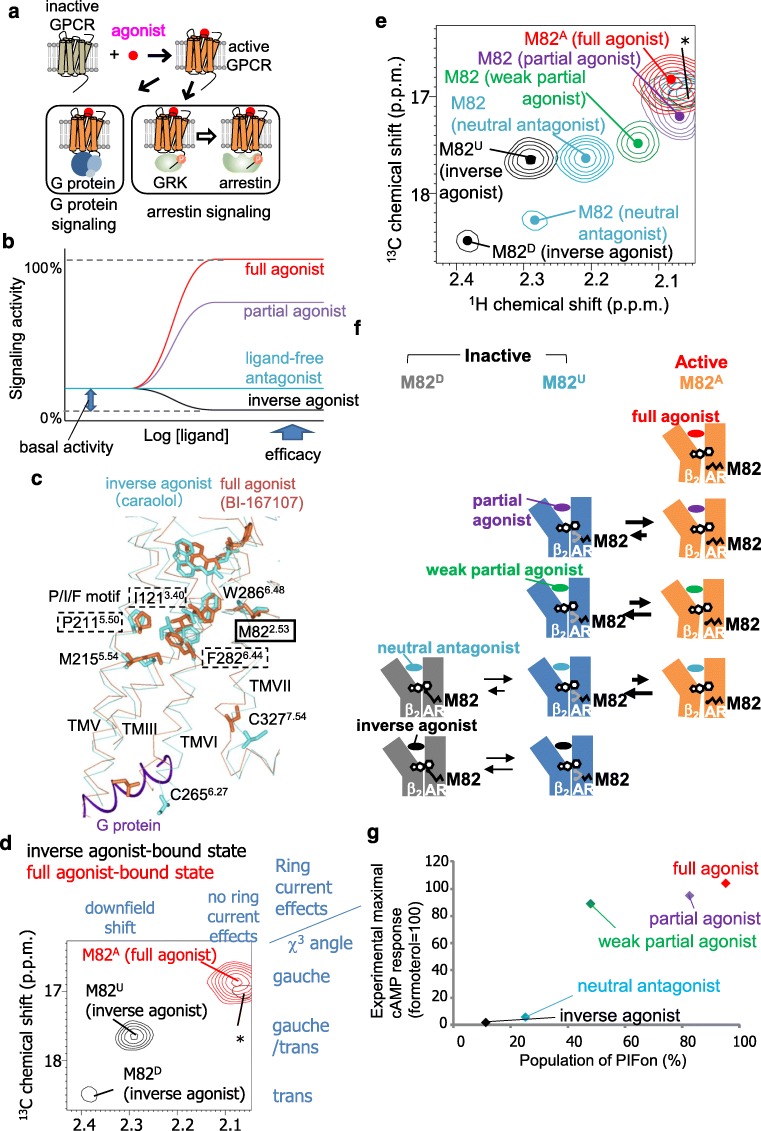
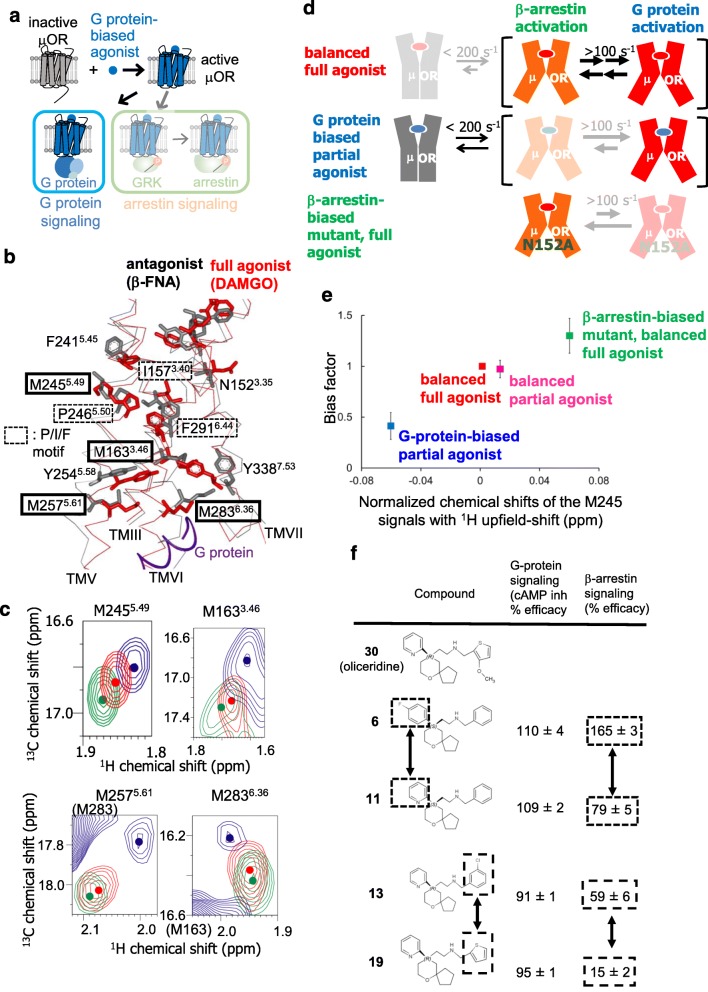
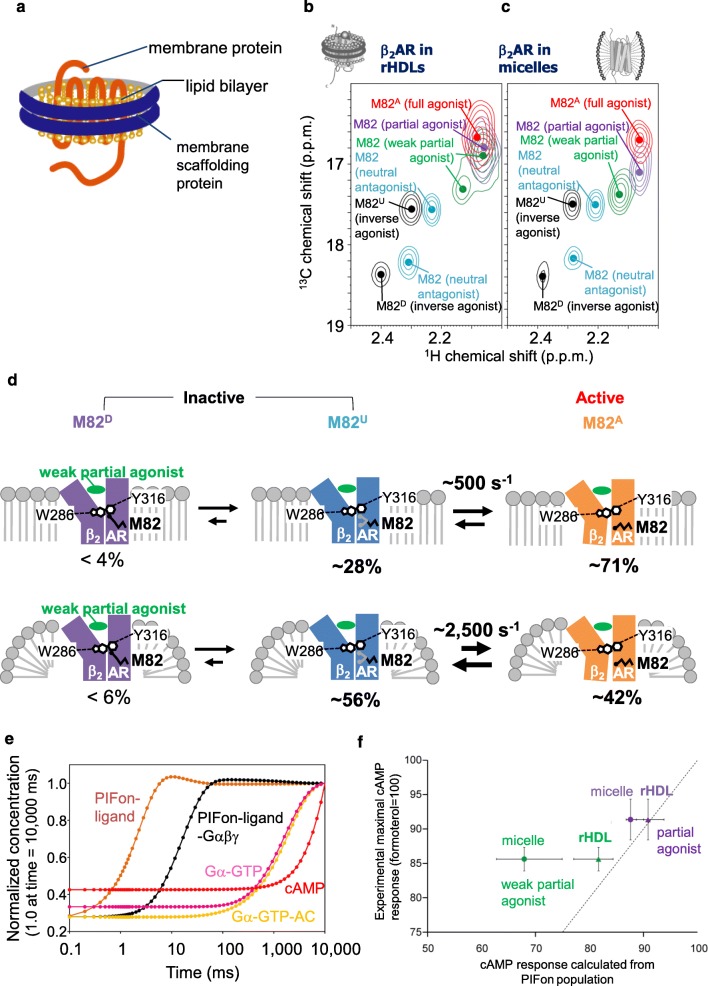
G protein-coupled receptors (GPCRs) function as receptors for various neurotransmitters, hormones, cytokines, and metabolites. GPCR ligands impart differing degrees of signaling in the G protein and arrestin pathways, in phenomena called biased signaling, and each ligand for a given GPCR has a characteristic level of ability to activate or deactivate its target, which is referred to as its efficacy. The ligand efficacies and biased signaling of GPCRs remarkably affect the therapeutic properties of the ligands. However, these features of GPCRs can only be partially understood from the crystallography data, although numerous GPCR structures have been solved. NMR analyses have revealed that GPCRs have multiple interconverting substates, exchanging on various timescales, and that the exchange rates are related to the ligand efficacies and biased signaling. In addition, NMR analyses of GPCRs in the lipid bilayer environment of rHDLs revealed that the exchange rates are modulated by the lipid bilayer environment, highlighting the importance of the function-related dynamics in the lipid bilayer. In this review, we will describe several solution NMR studies that have clarified the conformational dynamics related to the ligand efficacy and biased signaling of GPCRs.
Keywords: Adrenergic receptor; Membrane protein; Nanodiscs; Nuclear magnetic resonance; Opioid receptor.
Conflict of interest statement
Takumi Ueda declares that he has no conflict of interest. Yutaka Kofuku declares that he has no conflict of interest. Junya Okude declares that he has no conflict of interest. Shunsuke Imai declares that he has no conflict of interest. Yutaro Shiraishi declares that he has no conflict of interest. Ichio Shimada declares that he has no conflict of interest.
Figures
Similar articles
-
Identification of a Conformational Equilibrium That Determines the Efficacy and Functional Selectivity of the μ-Opioid Receptor.Angew Chem Int Ed Engl. 2015 Dec 21;54(52):15771-6. doi: 10.1002/anie.201508794. Epub 2015 Nov 16. Angew Chem Int Ed Engl. 2015. PMID: 26568421 Free PMC article.
-
Function-related dynamics of GPCRs.J Magn Reson. 2022 Mar;336:107164. doi: 10.1016/j.jmr.2022.107164. Epub 2022 Feb 7. J Magn Reson. 2022. PMID: 35168190
-
Functional dynamics of deuterated β2 -adrenergic receptor in lipid bilayers revealed by NMR spectroscopy.Angew Chem Int Ed Engl. 2014 Dec 1;53(49):13376-9. doi: 10.1002/anie.201406603. Epub 2014 Oct 3. Angew Chem Int Ed Engl. 2014. PMID: 25284766
-
Biased signaling of G protein coupled receptors (GPCRs): Molecular determinants of GPCR/transducer selectivity and therapeutic potential.Pharmacol Ther. 2019 Aug;200:148-178. doi: 10.1016/j.pharmthera.2019.05.006. Epub 2019 May 8. Pharmacol Ther. 2019. PMID: 31075355 Review.
-
The role of structural dynamics in GPCR-mediated signaling.FEBS J. 2021 Apr;288(8):2461-2489. doi: 10.1111/febs.15841. FEBS J. 2021. PMID: 33871923 Review.
Cited by
-
Capturing Peptide-GPCR Interactions and Their Dynamics.Molecules. 2020 Oct 15;25(20):4724. doi: 10.3390/molecules25204724. Molecules. 2020. PMID: 33076289 Free PMC article. Review.
-
Activation Microswitches in Adenosine Receptor A2A Function as Rheostats in the Cell Membrane.Biochemistry. 2020 Oct 27;59(42):4059-4071. doi: 10.1021/acs.biochem.0c00626. Epub 2020 Oct 15. Biochemistry. 2020. PMID: 33054162 Free PMC article.
-
Cellular lipids regulate the conformational ensembles of the disordered intracellular loop 3 in β2-adrenergic receptor.iScience. 2024 May 23;27(6):110086. doi: 10.1016/j.isci.2024.110086. eCollection 2024 Jun 21. iScience. 2024. PMID: 38947516 Free PMC article.
-
Opportunities and Challenges of Backbone, Sidechain, and RDC Experiments to Study Membrane Protein Dynamics in a Detergent-Free Lipid Environment Using Solution State NMR.Front Mol Biosci. 2019 Oct 25;6:103. doi: 10.3389/fmolb.2019.00103. eCollection 2019. Front Mol Biosci. 2019. PMID: 31709261 Free PMC article. Review.
-
Insights into the structure and activation mechanism of some class B1 GPCR family members.Mol Biol Rep. 2024 Sep 6;51(1):966. doi: 10.1007/s11033-024-09876-w. Mol Biol Rep. 2024. PMID: 39240462 Review.
References
-
- Ballesteros JA, Weinstein H. Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G-protein coupled receptors. Methods Neurosci. 1995;25:366–428.
-
- Bayburt TH, Grinkova YV, Sligar SG. Self-assembly of discoidal phospholipid bilayer nanoparticles with membrane scaffold proteins. Nano Lett. 2002;2:853–856.
-
- Bohn LM, Lefkowitz RJ, Gainetdinov RR, Peppel K, Caron MG, Lin FT. Enhanced morphine analgesia in mice lacking β-arrestin 2. Science. 1999;286:2495–2498. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
