

Reactive Enamines and Imines In Vivo: Lessons from the RidA Paradigm
- PMID: 31103411
- PMCID: PMC6760865
- DOI: 10.1016/j.tibs.2019.04.011
Reactive Enamines and Imines In Vivo: Lessons from the RidA Paradigm
Abstract
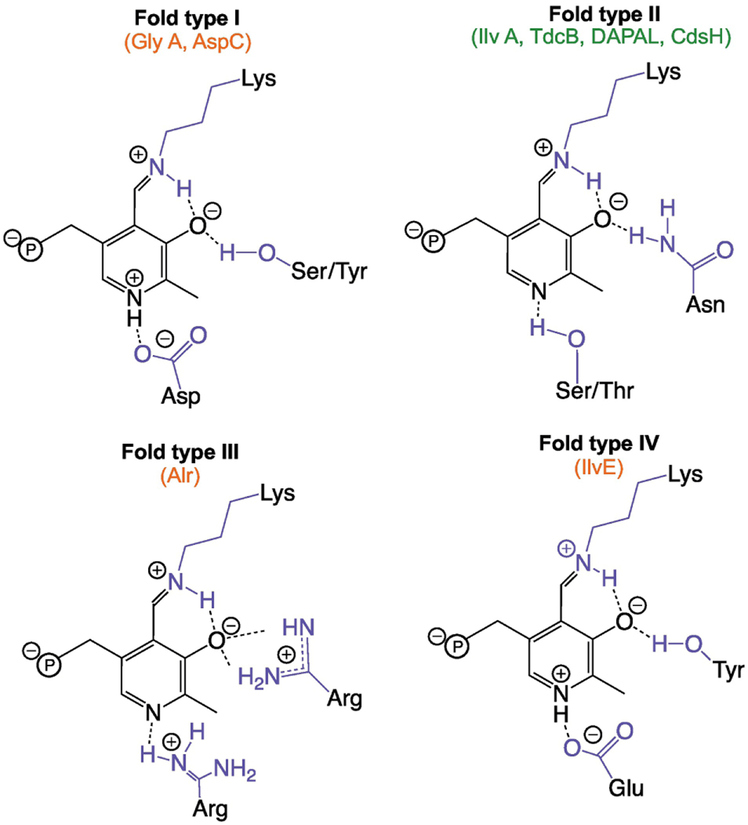
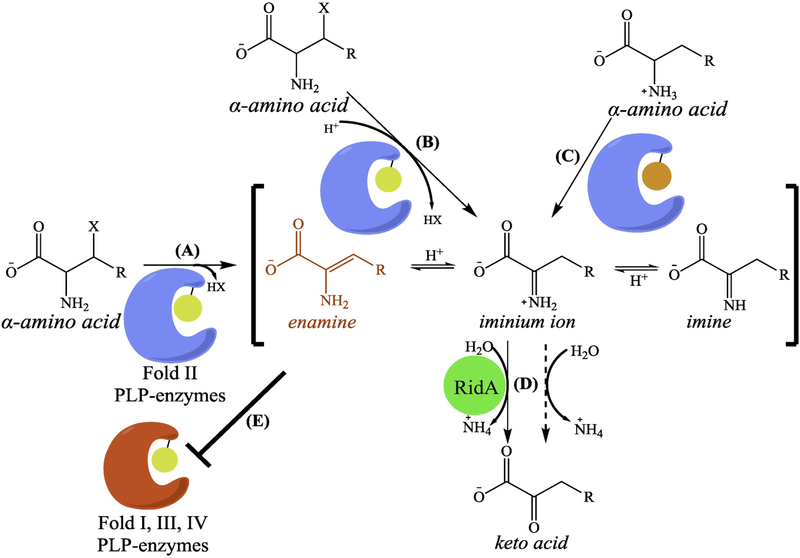
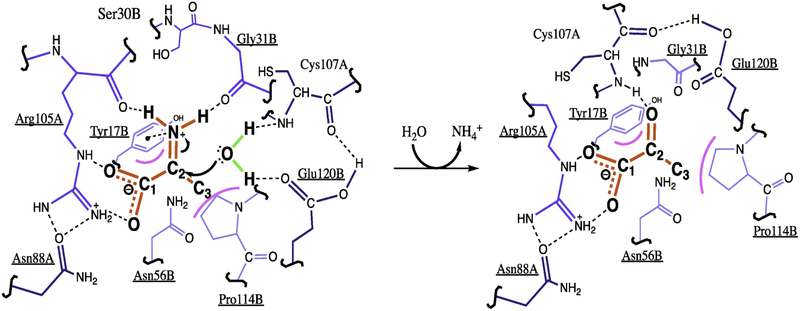
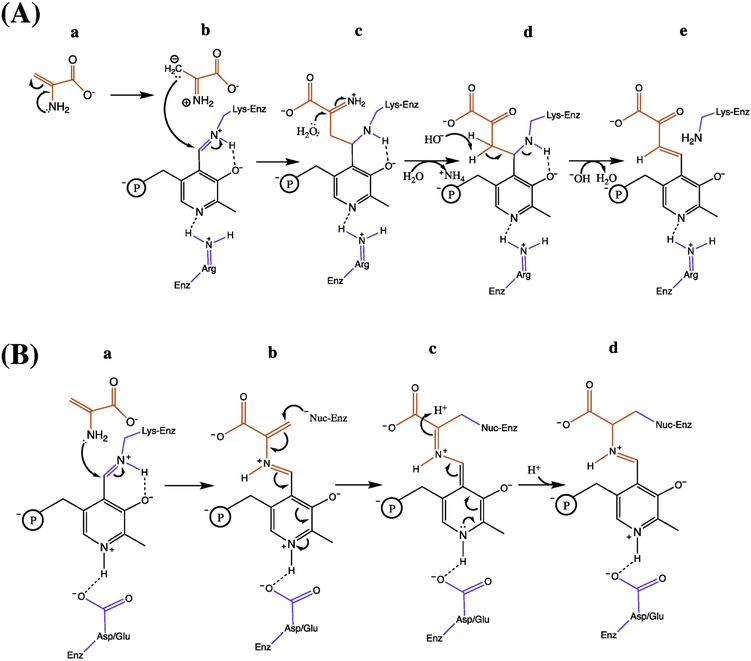
Metabolic networks are webs of integrated reactions organized to maximize growth and replication while minimizing the detrimental impact that reactive metabolites can have on fitness. Enamines and imines, such as 2-aminoacrylate (2AA), are reactive metabolites produced as short-lived intermediates in a number of enzymatic processes. Left unchecked, the inherent reactivity of enamines and imines may perturb the metabolic network. Genetic and biochemical studies have outlined a role for the broadly conserved reactive intermediate deaminase (Rid) (YjgF/YER057c/UK114) protein family, in particular RidA, in catalyzing the hydrolysis of enamines and imines to their ketone product. Herein, we discuss new findings regarding the biological significance of enamine and imine production and outline the importance of RidA in controlling the accumulation of reactive metabolites.
Keywords: 2-aminoacrylate stress; RidA; enamine/imine metabolism; reactive metabolite.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
RidA Proteins Protect against Metabolic Damage by Reactive Intermediates.Microbiol Mol Biol Rev. 2020 Jul 15;84(3):e00024-20. doi: 10.1128/MMBR.00024-20. Print 2020 Aug 19. Microbiol Mol Biol Rev. 2020. PMID: 32669283 Free PMC article. Review.
-
Genomic and experimental evidence for multiple metabolic functions in the RidA/YjgF/YER057c/UK114 (Rid) protein family.BMC Genomics. 2015 May 15;16(1):382. doi: 10.1186/s12864-015-1584-3. BMC Genomics. 2015. PMID: 25975565 Free PMC article.
-
Evidence for a Negative Correlation between Human Reactive Enamine-Imine Intermediate Deaminase A (RIDA) Activity and Cell Proliferation Rate: Role of Lysine Succinylation of RIDA.Int J Mol Sci. 2021 Apr 7;22(8):3804. doi: 10.3390/ijms22083804. Int J Mol Sci. 2021. PMID: 33916919 Free PMC article.
-
RidA proteins prevent metabolic damage inflicted by PLP-dependent dehydratases in all domains of life.mBio. 2013 Feb 5;4(1):e00033-13. doi: 10.1128/mBio.00033-13. mBio. 2013. PMID: 23386433 Free PMC article.
-
From microbiology to cancer biology: the Rid protein family prevents cellular damage caused by endogenously generated reactive nitrogen species.Mol Microbiol. 2015 Apr;96(2):211-9. doi: 10.1111/mmi.12945. Epub 2015 Feb 26. Mol Microbiol. 2015. PMID: 25620221 Free PMC article. Review.
Cited by
-
Site-directed mutagenesis reveals the interplay between stability, structure, and enzymatic activity in RidA from Capra hircus.Protein Sci. 2024 Jun;33(6):e5036. doi: 10.1002/pro.5036. Protein Sci. 2024. PMID: 38801230 Free PMC article.
-
Serendipity Reveals the Function and Physiological Role of a Large Family of Proteins.J Bacteriol. 2022 Feb 15;204(2):e0055621. doi: 10.1128/JB.00556-21. Epub 2021 Dec 6. J Bacteriol. 2022. PMID: 34871032 Free PMC article.
-
His-163 is a stereospecific proton donor in the mechanism of d-glucosaminate-6-phosphate ammonia-lyase.FEBS Lett. 2022 Sep;596(18):2441-2448. doi: 10.1002/1873-3468.14469. Epub 2022 Aug 29. FEBS Lett. 2022. PMID: 35953460 Free PMC article.
-
Chemical proteomics enhances the understanding of 2AA stress in Salmonella enterica.mSystems. 2025 Jun 17;10(6):e0054025. doi: 10.1128/msystems.00540-25. Epub 2025 May 29. mSystems. 2025. PMID: 40439409 Free PMC article.
-
Tetrahydrofolate levels influence 2-aminoacrylate stress in Salmonella enterica.J Bacteriol. 2024 Apr 18;206(4):e0004224. doi: 10.1128/jb.00042-24. Epub 2024 Apr 2. J Bacteriol. 2024. PMID: 38563759 Free PMC article.
References
-
- de Lorenzo V et al. (2015) Chemical reactivity drives spatiotemporal organisation of bacterial metabolism. FEMS Microbiol. Rev 39 (1), 96–119. - PubMed
-
- Albert R et al. (2000) Error and attack tolerance of complex networks. Nature 406 (6794), 378–382. - PubMed
-
- de Crecy-Lagard V et al. (2018) Newly-discovered enzymes that function in metabolite damage-control. Curr. Opin. Chem. Biol 47, 101–108. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
