

Influenza Antigen Engineering Focuses Immune Responses to a Subdominant but Broadly Protective Viral Epitope
- PMID: 31104946
- PMCID: PMC6748655
- DOI: 10.1016/j.chom.2019.04.003
Influenza Antigen Engineering Focuses Immune Responses to a Subdominant but Broadly Protective Viral Epitope
Abstract
Viral glycoproteins are under constant immune surveillance by a host's adaptive immune responses. Antigenic variation including glycan introduction or removal is among the mechanisms viruses have evolved to escape host immunity. Understanding how glycosylation affects immunodominance on complex protein antigens may help decipher underlying B cell biology. To determine how B cell responses can be altered by such modifications, we engineered glycans onto the influenza virus hemagglutinin (HA) and characterized the molecular features of the elicited humoral immunity in mice. We found that glycan addition changed the initially diverse antibody repertoire into an epitope-focused, genetically restricted response. Structural analyses showed that one antibody gene family targeted a previously subdominant, occluded epitope at the head interface. Passive transfer of this antibody conferred Fc-dependent protection to influenza virus-challenged mice. These results have potential implications for next-generation viral vaccines aimed at directing B cell responses to preferred epitope(s).
Keywords: broadly neutralizing antibodies; immunogen design; influenza hemagglutinin; protein engineering.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
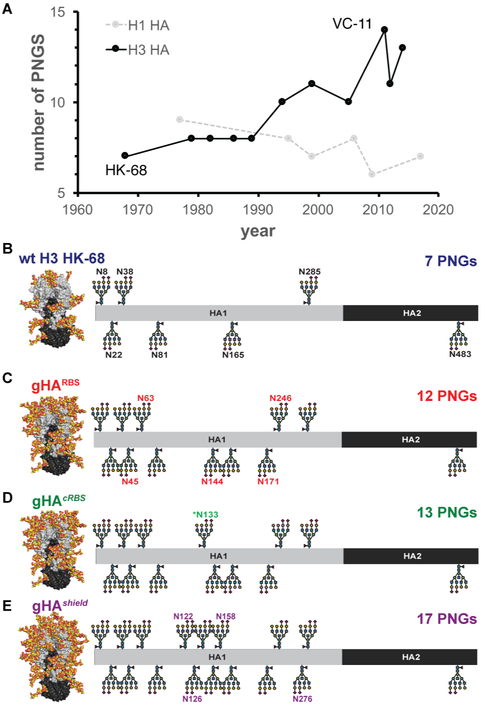
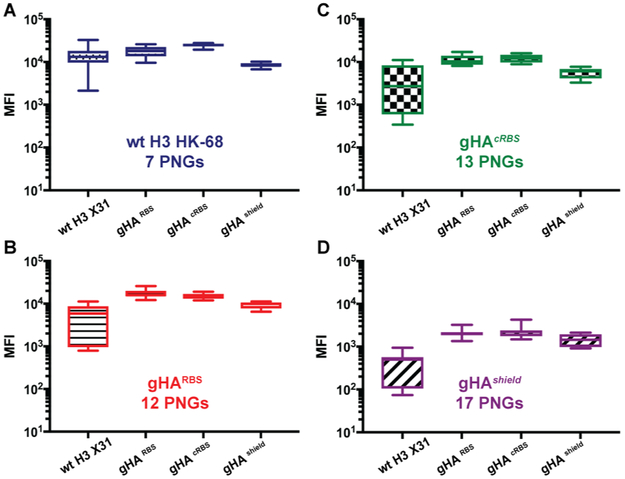
Comment in
-
"Breathing" Hemagglutinin Reveals Cryptic Epitopes for Universal Influenza Vaccine Design.Cell. 2019 May 16;177(5):1086-1088. doi: 10.1016/j.cell.2019.04.034. Cell. 2019. PMID: 31100263 Free PMC article.
References
-
- Bizebard T, Gigant B, Rigolet P, Rasmussen B, Diat O, Bosecke P, Wharton SA, Skehel JJ, and Knossow M (1995). Structure of influenza virus haemagglutinin complexed with a neutralizing antibody. Nature 376, 92–94. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
