

RNA-seq data reveals a coordinated regulation mechanism of multigenes involved in the high accumulation of palmitoleic acid and oil in sea buckthorn berry pulp
- PMID: 31109294
- PMCID: PMC6528223
- DOI: 10.1186/s12870-019-1815-x
RNA-seq data reveals a coordinated regulation mechanism of multigenes involved in the high accumulation of palmitoleic acid and oil in sea buckthorn berry pulp
Abstract
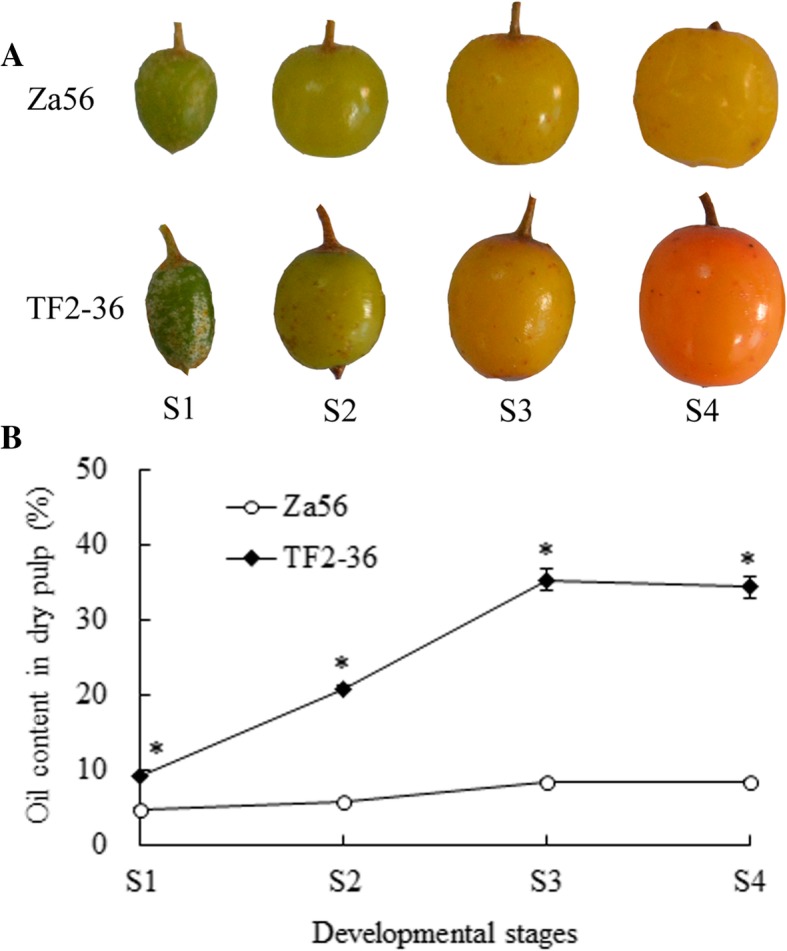
Background: Sea buckthorn is a woody oil crop in which palmitoleic acid (C16:1n7, an omega-7 fatty acid (FA)) contributes approximately 40% of the total FA content in berry pulp (non-seed tissue). However, the molecular mechanisms contributing to the high accumulation of C16:1n7 in developing sea buckthorn berry pulp (SBP) remain poorly understood.
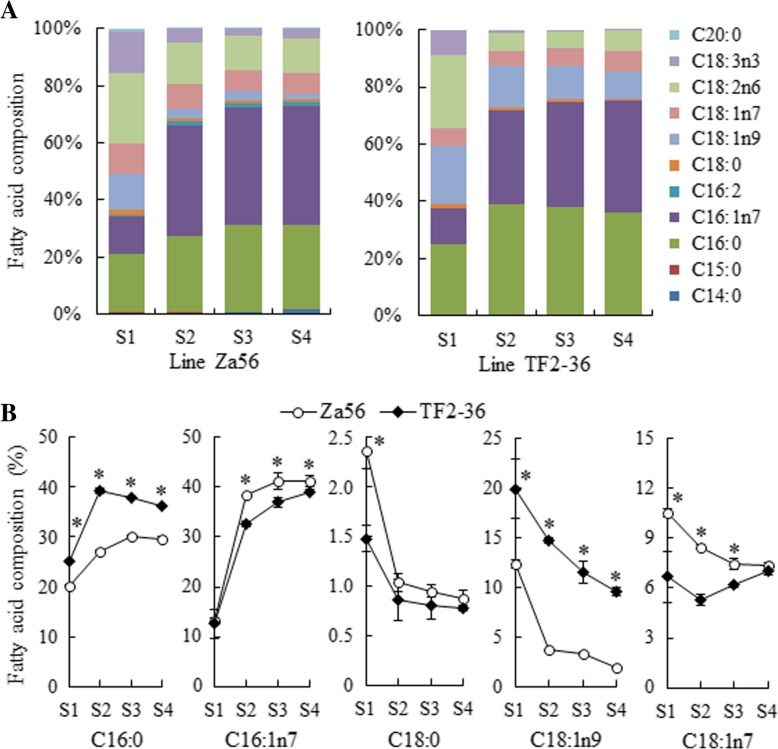
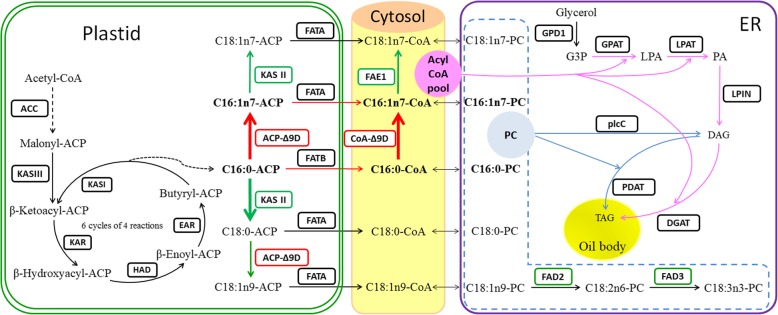
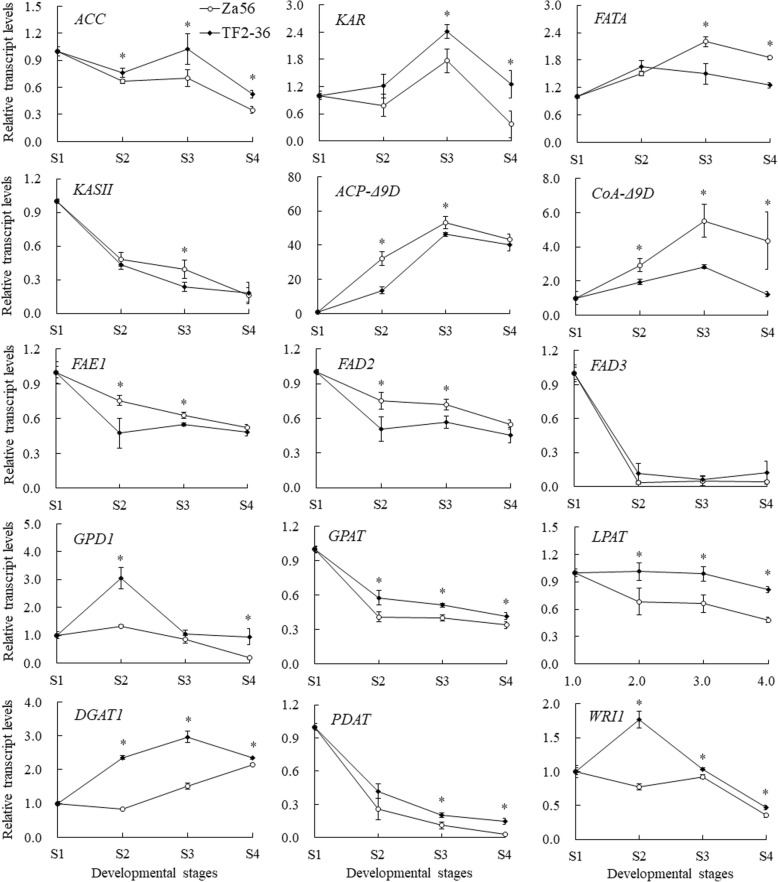
Results: We identified 1737 unigenes associated with lipid metabolism through RNA-sequencing analysis of the four developmental stages of berry pulp in two sea buckthorn lines, 'Za56' and 'TF2-36'; 139 differentially expressed genes were detected between the different berry pulp developmental stages in the two lines. Analyses of the FA composition showed that the C16:1n7 contents were significantly higher in line 'Za56' than in line 'TF2-36' in the mid-late developmental stages of SBP. Additionally, qRT-PCR analyses of 15 genes involved in FA and triacylglycerol (TAG) biosynthesis in both lines revealed that delta9-ACP-desaturase (ACP-Δ9D) competed with 3-ketoacyl-ACP-synthase II (KASII) for the substrate C16:0-ACP and that ACP-Δ9D and delta9-CoA-desaturase (CoA-Δ9D) gene expression positively correlated with C16:1n7 content; KASII and fatty acid elongation 1 (FAE1) gene expression positively correlated with C18:0 content in developing SBP. Specifically, the abundance of ACP-Δ9D and CoA-Δ9D transcripts in line 'Za56', which had a higher C16:1n7 content than line 'TF2-36', suggests that these two genes play an important role in C16:1n7 biosynthesis. Furthermore, the high expressions of the glycerol-3-phosphate dehydrogenase (GPD1) gene and the WRINKLED1 (WRI1) transcription factor contributed to increased biosynthesis of TAG precursor and FAs, respectively, in the early developmental stages of SBP, and the high expression of the diacylglycerol O-acyltransferase 1 (DGAT1) gene increased TAG assembly in the later developmental stages of SBP. Overall, we concluded that increased ACP-Δ9D and CoA-Δ9D levels coupled with decreased KASII and FAE1 activity is a critical event for high C16:1n7 accumulation and that the coordinated high expression of WRI1, GPD1, and DGAT1 genes resulted in high oil accumulation in SBP.
Conclusion: Our results provide a scientific basis for understanding the mechanism of high C16:1n7 accumulation in berry pulp (non-seed tissue) and are valuable to the genetic breeding programme for achieving a high quality and yield of SBP oil.
Keywords: Berry pulp oil; Fatty acid biosynthesis; Hippophae L.; Non-seed tissue; Oil accumulation; Palmitoleic acid.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Ruan CJ, Rumpunen K, Nybom H. Advances in improvement of quality and resistance in a multipurpose crop: sea buckthorn. Crit Rev Biotechnol. 2013;33(2):126–144. - PubMed
-
- Ding J, Ruan CJ, Guan Y, Shan JY. Comparison of oil contents and fatty acid composition between developing sea buckthorn pulp and seed. China Oil and Fats. 2017;42(5):140–144.
-
- Yang B, Kallio H. Fatty acid composition of lipids in sea buckthorn (Hippophae rhamnoides L.) berries of different origins. J Agric Food Chem. 2001;49(4):1939–1947. - PubMed
-
- Li W, Ruan CJ, Teixeira da Silva JA, Guo H, Zhao CE. NMR metabolomics of berry quality in sea buckthorn (Hippophae L.) Mol Breeding. 2013;31(1):57–67.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
