

Physiological responses and proteomic changes reveal insights into Stylosanthes response to manganese toxicity
- PMID: 31113380
- PMCID: PMC6530018
- DOI: 10.1186/s12870-019-1822-y
Physiological responses and proteomic changes reveal insights into Stylosanthes response to manganese toxicity
Abstract
Background: Manganese (Mn), an essential element for plants, can be toxic when present in excess. Stylo (Stylosanthes) is a pioneer tropical legume with great potential for Mn tolerance, but its Mn tolerance mechanisms remain poorly understood.
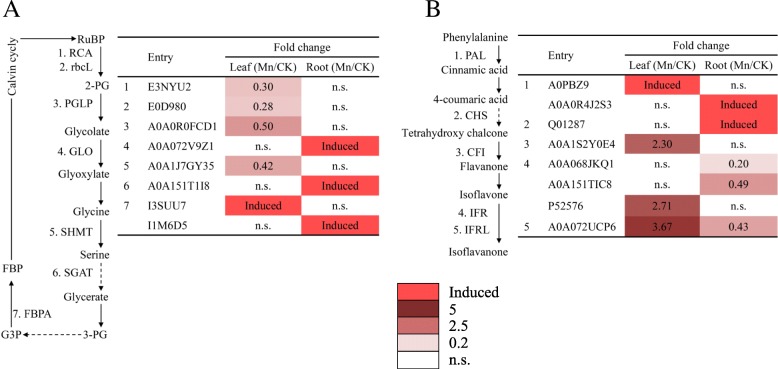
Results: In this study, variations in Mn tolerance were observed among nine stylo genotypes. Stylo genotype 'RY5' exhibited the highest Mn tolerance compared to the other tested genotypes, whereas 'TF2001' was a Mn-sensitive genotype. The mechanisms underlying the response of stylo to Mn toxicity were further investigated using these two genotypes with contrasting Mn tolerance. Results showed that stylo genotype RY5 exhibited Mn tolerance superior to that of genotype TF2001, showing lower reductions in leaf chlorophyll concentration, chlorophyll fluorescence parameters, photosynthetic indexes and plant dry weight under Mn toxicity. A label-free quantitative proteomic analysis was conducted to investigate the protein profiles in the leaves and roots of RY5 in response to Mn toxicity. A total of 356 differentially expressed proteins (DEPs) were identified, including 206 proteins from leaves and 150 proteins from roots, which consisted of 71 upregulated, 62 downregulated, 127 strongly induced and 96 completely suppressed proteins. These DEPs were mainly involved in defense response, photosynthesis, carbon fixation, metabolism, cell wall modulation and signaling. The qRT-PCR analysis verified that 10 out of 12 corresponding gene transcription patterns correlated with their encoding proteins after Mn exposure. Finally, a schematic was constructed to reveal insights into the molecular processes in the leaves and roots of stylo in response to Mn toxicity.
Conclusions: These findings suggest that stylo plants may cope with Mn toxicity by enhancing their defense response and phenylpropanoid pathways, adjusting photosynthesis and metabolic processes, and modulating protein synthesis and turnover. This study provides a platform for the future study of Mn tolerance mechanisms in stylo and may lead to a better understanding of the potential mechanisms underlying tropical legume adaptation to Mn toxicity.
Keywords: Antioxidant protection; Manganese toxicity; Oxidative stress; Proteomics; Stylosanthes.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures








References
-
- Sparrow LA, Uren NC. Manganese oxidation and reduction in soils: effects of temperature, water potential, pH and their interactions. Soil Res. 2014;52(5):483–494. doi: 10.1071/SR13159. - DOI
-
- Nagajyoti PC, Lee KD, Sreekanth TVM. Heavy metals, occurrence and toxicity for plants: a review. Environ Chem Lett. 2010;8(3):199–216. doi: 10.1007/s10311-010-0297-8. - DOI
-
- Millaleo R, Reyes-Díaz M, Ivanov AG, Mora ML, Alberdi M. Manganese as essential and toxic element for plants: transport, accumulation and resistance mechanism. J Soil Sci Plant Nutr. 2010;10(4):470–481. doi: 10.4067/S0718-95162010000200008. - DOI
MeSH terms
Substances
Grants and funding
- 31801951, 31672483, 31861143013/National Natural Science Foundation of China
- QCXM201715/Young Talents Academic Innovation Project of Hainan Association for Science and Technology
- ZDYF2018048/Key Research and Development Program of Hainan
- CARS-34/Modern Agro-industry Technology Research System
- 1630032018004/Central Public-interest Scientific Institution Basal Research Fund for CATAS
LinkOut - more resources
Full Text Sources
