

mTORC1 amplifies the ATF4-dependent de novo serine-glycine pathway to supply glycine during TGF-β1-induced collagen biosynthesis
- PMID: 31113850
- PMCID: PMC6584619
- DOI: 10.1126/scisignal.aav3048
mTORC1 amplifies the ATF4-dependent de novo serine-glycine pathway to supply glycine during TGF-β1-induced collagen biosynthesis
Abstract
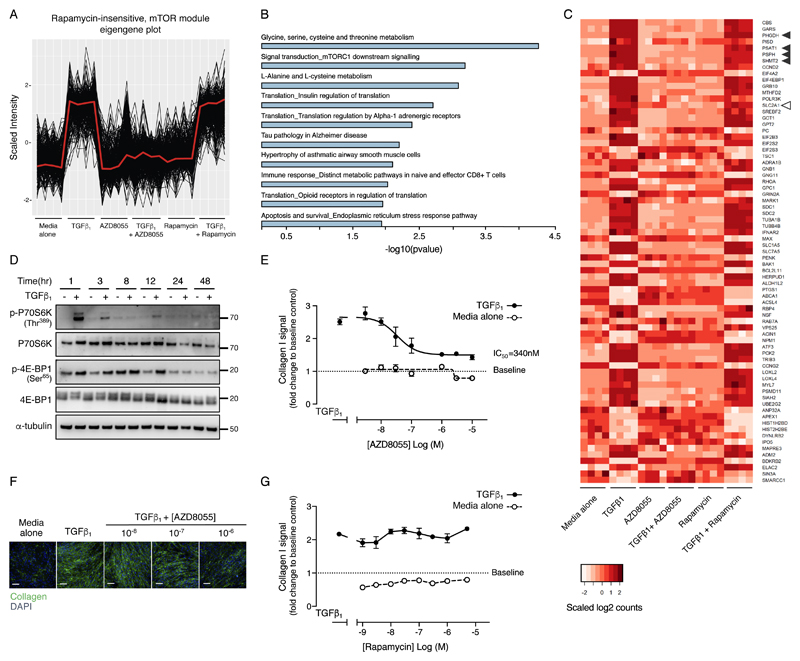
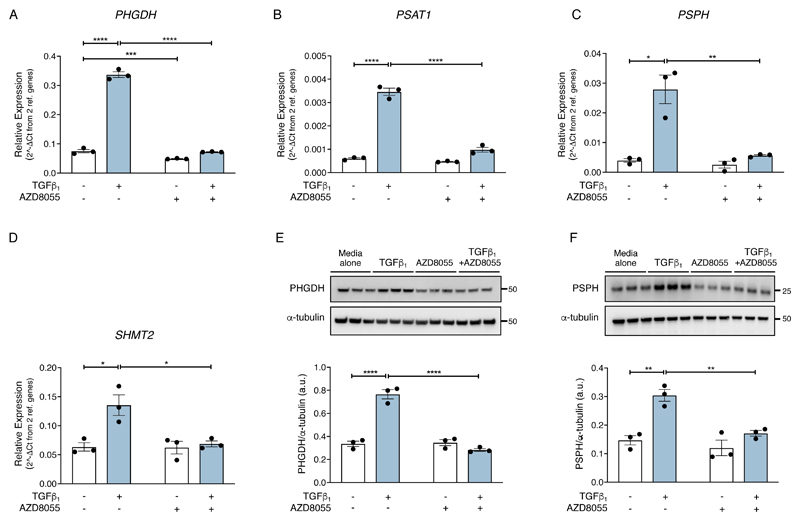
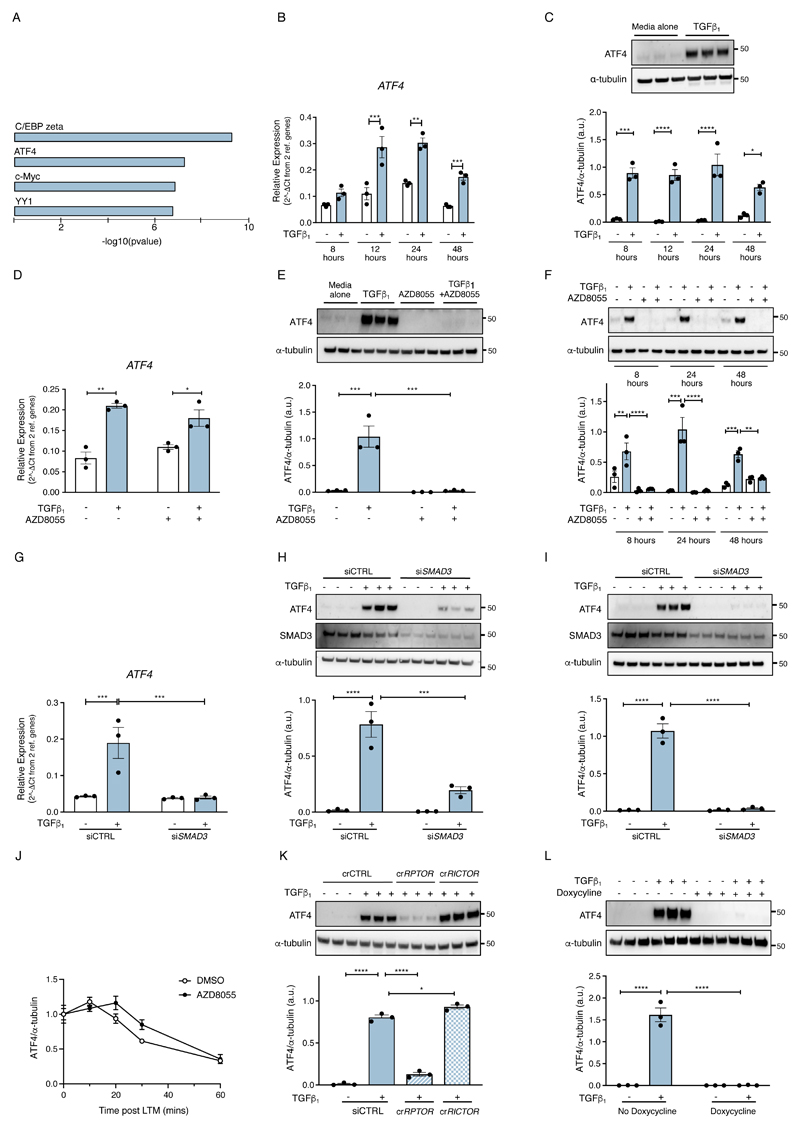
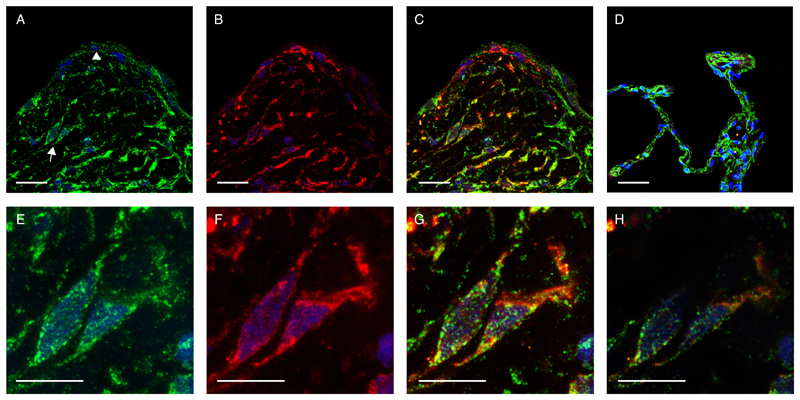
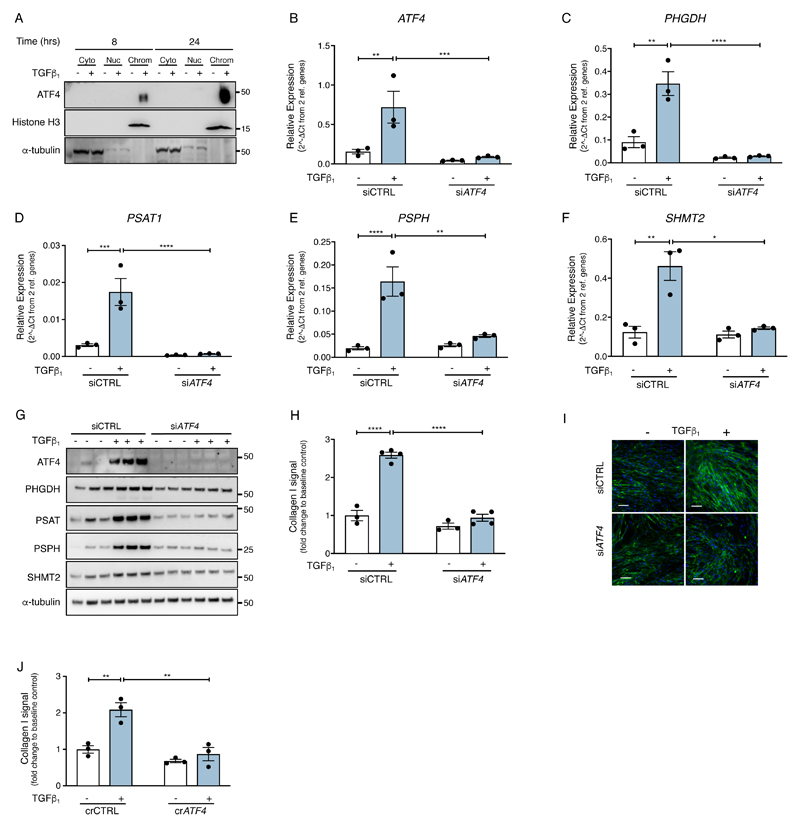
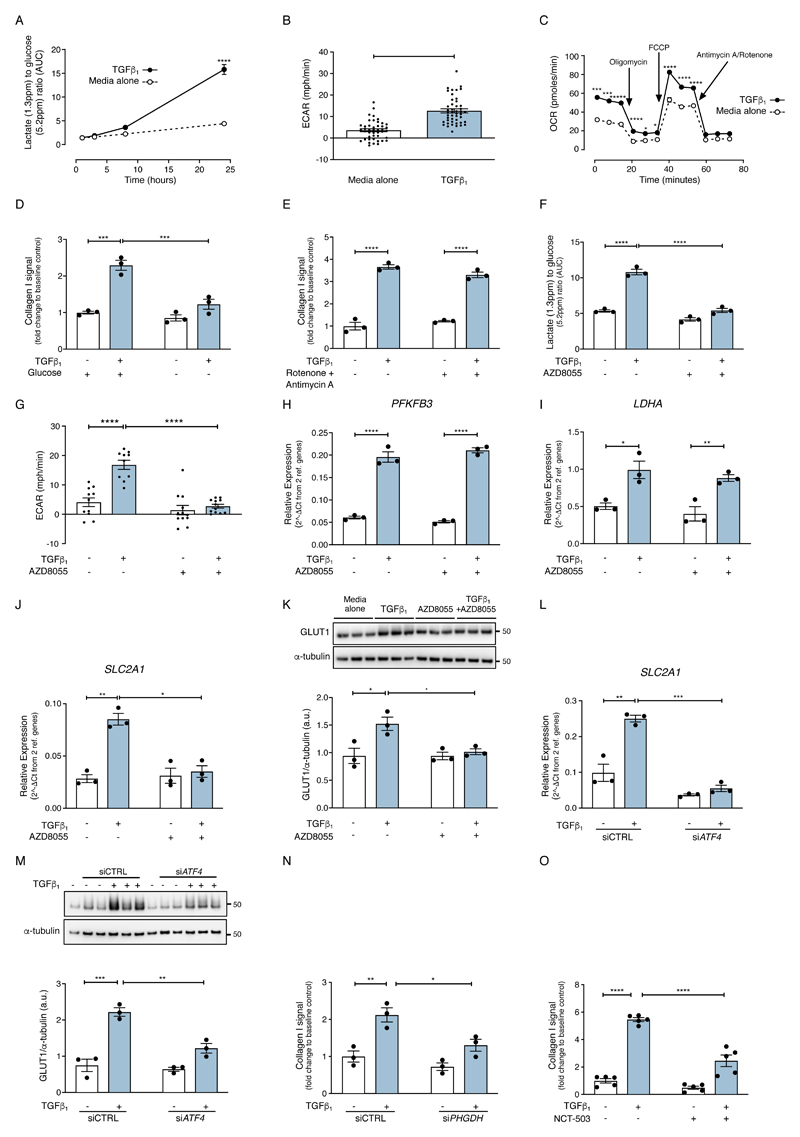
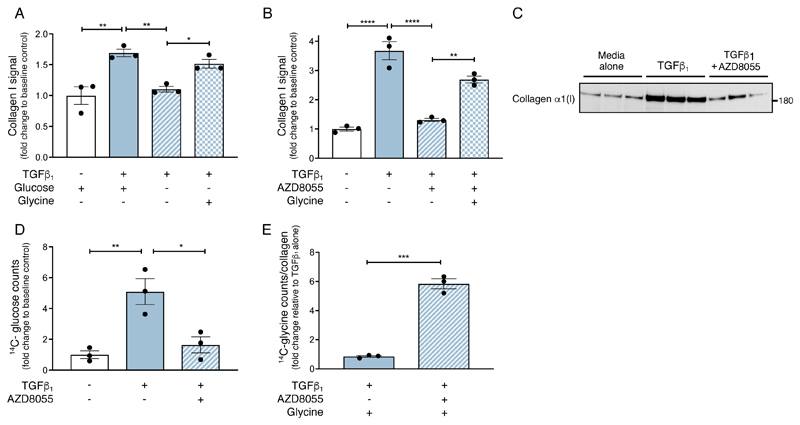
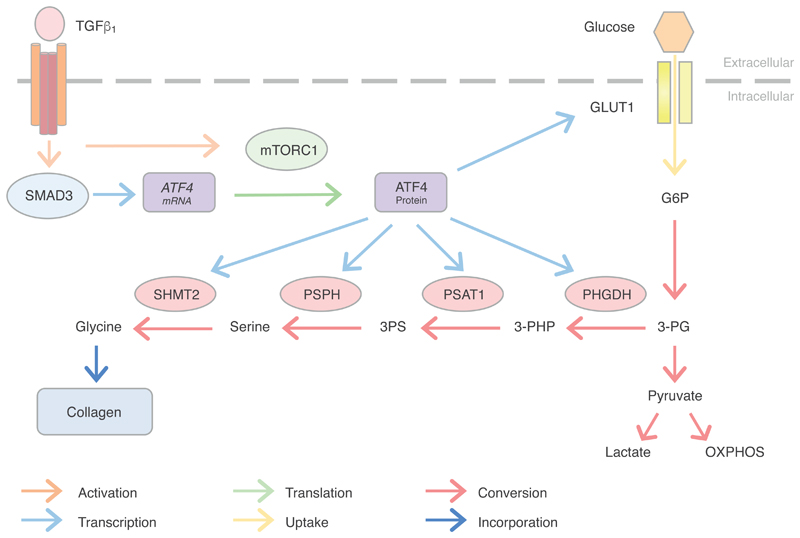
The differentiation of fibroblasts into a transient population of highly activated, extracellular matrix (ECM)-producing myofibroblasts at sites of tissue injury is critical for normal tissue repair. Excessive myofibroblast accumulation and persistence, often as a result of a failure to undergo apoptosis when tissue repair is complete, lead to pathological fibrosis and are also features of the stromal response in cancer. Myofibroblast differentiation is accompanied by changes in cellular metabolism, including increased glycolysis, to meet the biosynthetic demands of enhanced ECM production. Here, we showed that transforming growth factor-β1 (TGF-β1), the key pro-fibrotic cytokine implicated in multiple fibrotic conditions, increased the production of activating transcription factor 4 (ATF4), the transcriptional master regulator of amino acid metabolism, to supply glucose-derived glycine to meet the amino acid requirements associated with enhanced collagen production in response to myofibroblast differentiation. We further delineated the signaling pathways involved and showed that TGF-β1-induced ATF4 production depended on cooperation between canonical TGF-β1 signaling through Smad3 and activation of mechanistic target of rapamycin complex 1 (mTORC1) and its downstream target eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1). ATF4, in turn, promoted the transcription of genes encoding enzymes of the de novo serine-glycine biosynthetic pathway and glucose transporter 1 (GLUT1). Our findings suggest that targeting the TGF-β1-mTORC1-ATF4 axis may represent a novel therapeutic strategy for interfering with myofibroblast function in fibrosis and potentially in other conditions, including cancer.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Coultas DB, Zumwalt RE, Black WC, Sobonya RE. The epidemiology of interstitial lung diseases. Am J Respir Crit Care Med. 1994;150:967–72. - PubMed
-
- King TE, et al. Idiopathic pulmonary fibrosis: relationship between histopathologic features and mortality. Am J Respir Crit Care Med. 2001;164:1025–32. - PubMed
-
- Richeldi L, et al. Efficacy and safety of nintedanib in idiopathic pulmonary fibrosis. N Engl J Med. 2014;370:2071–82. - PubMed
-
- King TE, et al. A phase 3 trial of pirfenidone in patients with idiopathic pulmonary fibrosis. N Engl J Med. 2014;370:2083–92. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
