

Cerebellar Lobulus Simplex and Crus I Differentially Represent Phase and Phase Difference of Prefrontal Cortical and Hippocampal Oscillations
- PMID: 31116979
- PMCID: PMC6538275
- DOI: 10.1016/j.celrep.2019.04.085
Cerebellar Lobulus Simplex and Crus I Differentially Represent Phase and Phase Difference of Prefrontal Cortical and Hippocampal Oscillations
Abstract
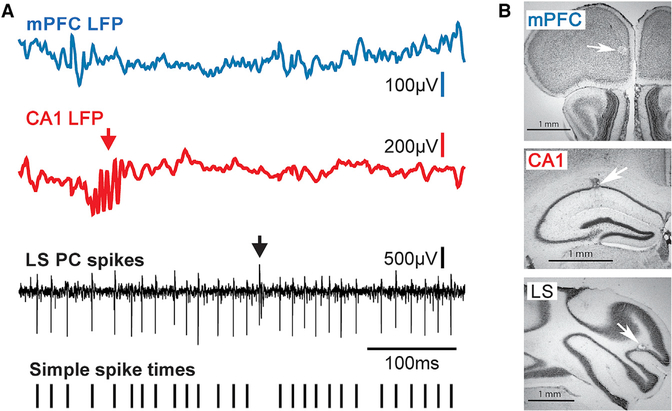
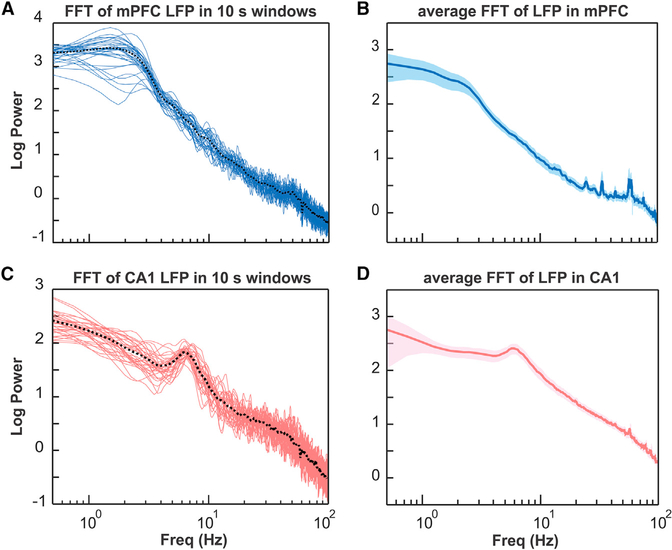
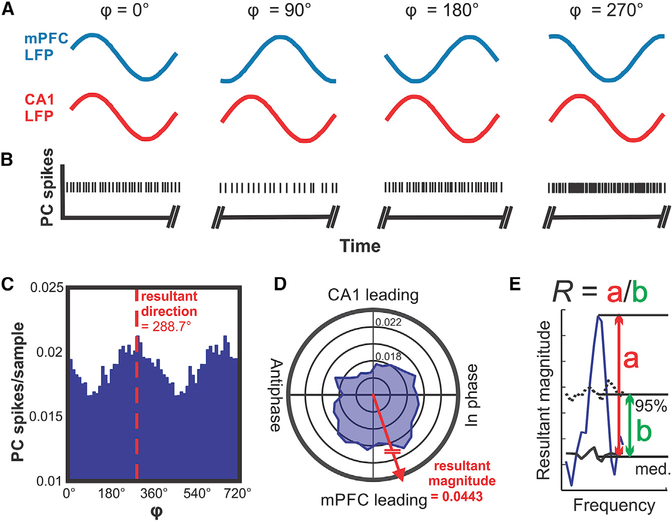
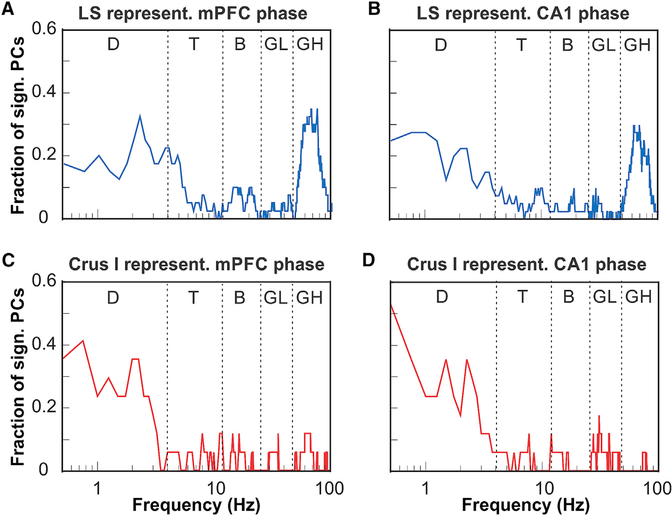
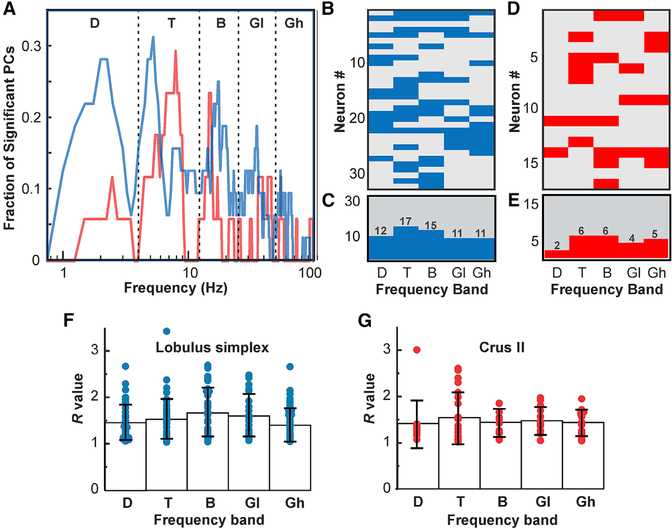
The cerebellum has long been implicated in tasks involving precise temporal control, especially in the coordination of movements. Here we asked whether the cerebellum represents temporal aspects of oscillatory neuronal activity, measured as instantaneous phase and difference between instantaneous phases of oscillations in two cerebral cortical areas involved in cognitive function. We simultaneously recorded Purkinje cell (PC) single-unit spike activity in cerebellar lobulus simplex (LS) and Crus I and local field potential (LFP) activity in the medial prefrontal cortex (mPFC) and dorsal hippocampus CA1 region (dCA1). Purkinje cells in cerebellar LS and Crus I differentially represented specific phases and phase differences of mPFC and dCA1 LFP oscillations in a frequency-specific manner, suggesting a site- and frequency-specific cerebellar representation of temporal aspects of neuronal oscillations in non-motor cerebral cortical areas. These findings suggest that cerebellar interactions with cerebral cortical areas involved in cognitive functions might involve temporal coordination of neuronal oscillations.
Keywords: cerebro-cerebellar interaction; neuronal oscillation; oscillation phase; phase difference.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures

References
-
- Bastos AM, Vezoli J, and Fries P (2015). Communication through coherence with inter-areal delays. Curr. Opin. Neurobiol 31, 173–180. - PubMed
-
- Braitenberg V (1961). Functional Interpretation of Cerebellar Histology. Nature 190, 539–540.
-
- Braitenberg V (1967). Is the Cerebellar Cortex a Biological Clock in the Millisecond Range? Prog. Brain Res 25, 334–346. - PubMed
-
- Braitenberg V, Heck D, and Sultan F (1997). The detection and generation of sequences as a key to cerebellar function: experiments and theory. Behav. Brain Sci 20, 229–245, discussion 245–277. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
