

Dissociable dopamine dynamics for learning and motivation
- PMID: 31118513
- PMCID: PMC6555489
- DOI: 10.1038/s41586-019-1235-y
Dissociable dopamine dynamics for learning and motivation
Erratum in
-
Publisher Correction: Dissociable dopamine dynamics for learning and motivation.Nature. 2019 Jul;571(7763):E3. doi: 10.1038/s41586-019-1353-6. Nature. 2019. PMID: 31217588
Abstract
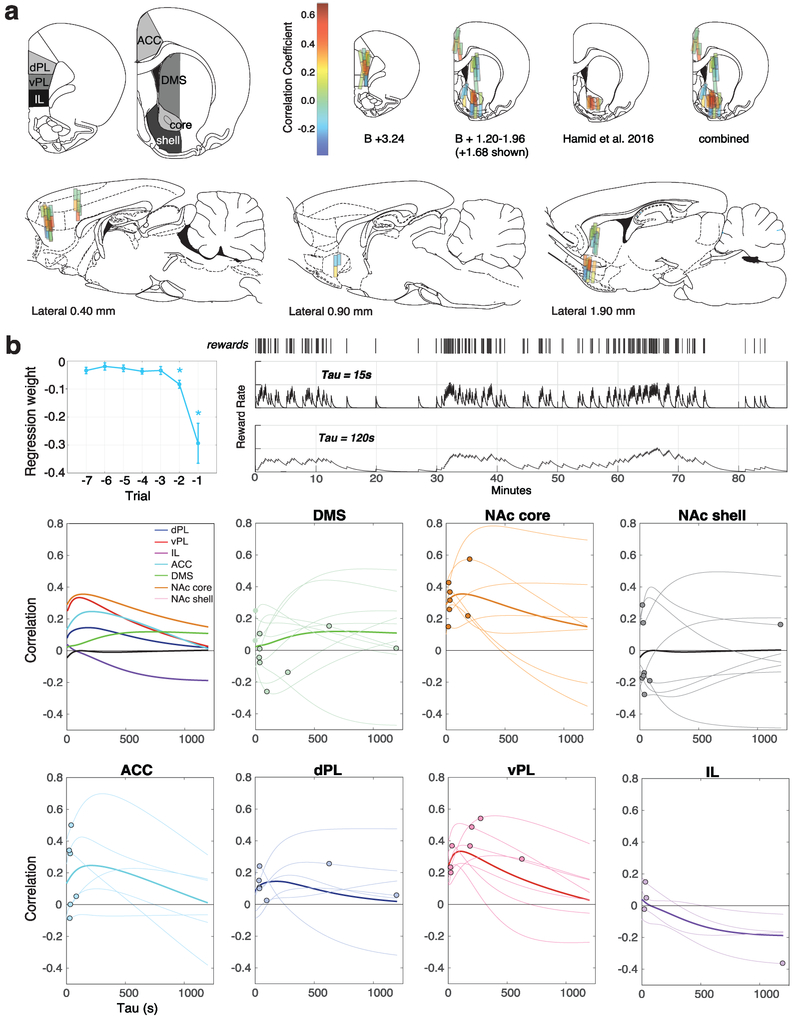
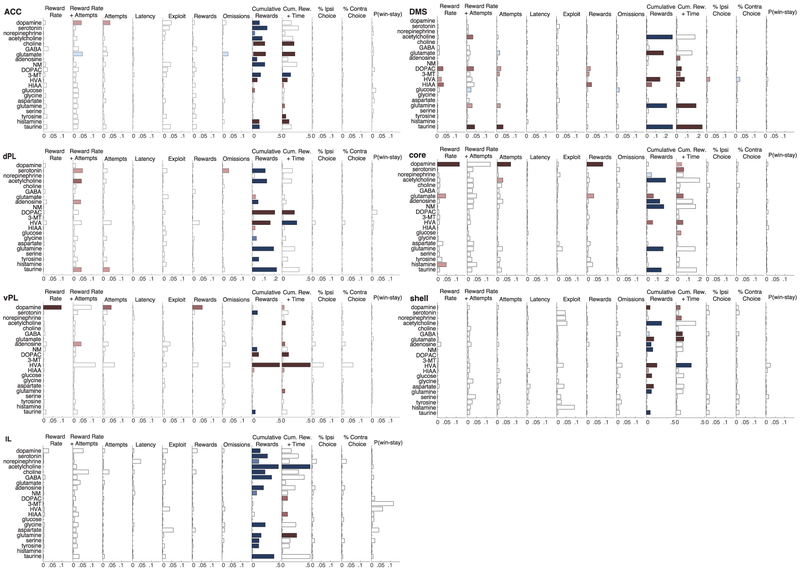
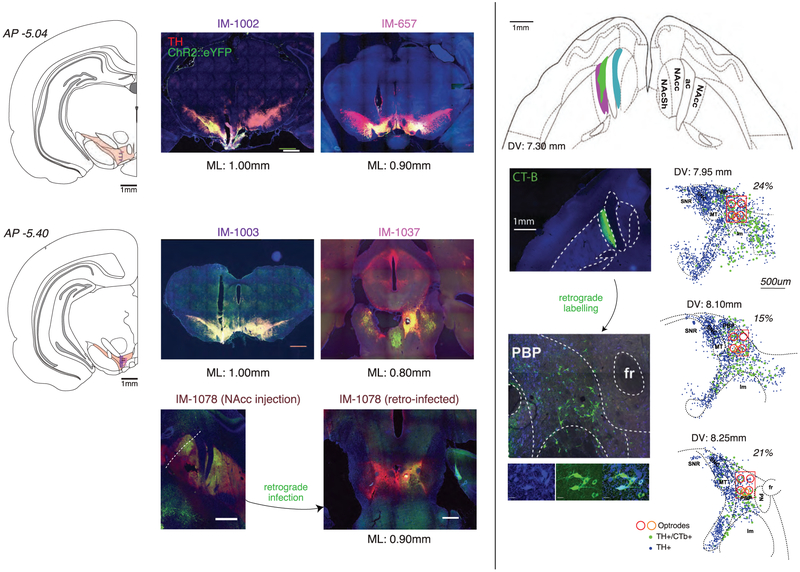
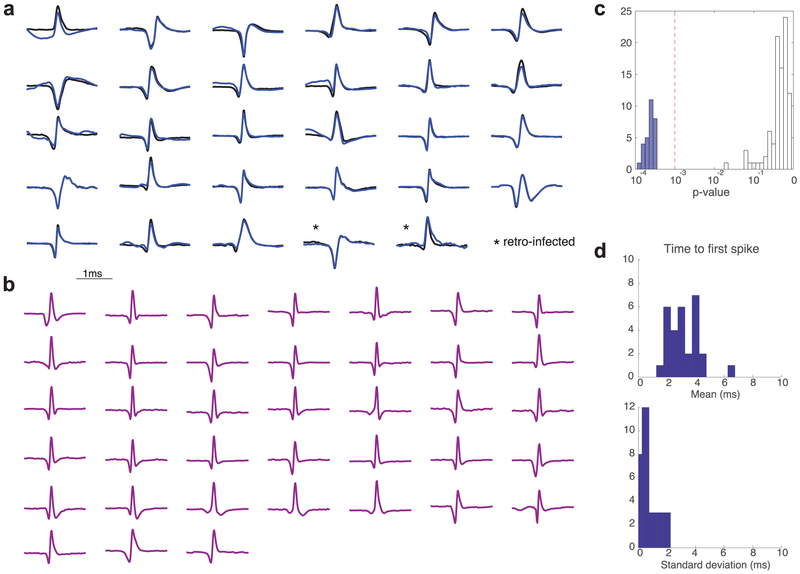
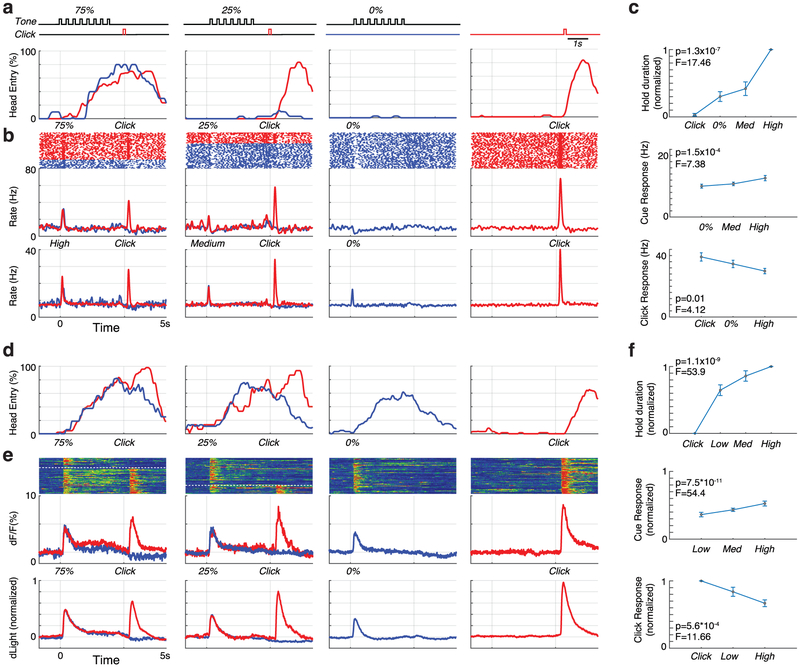
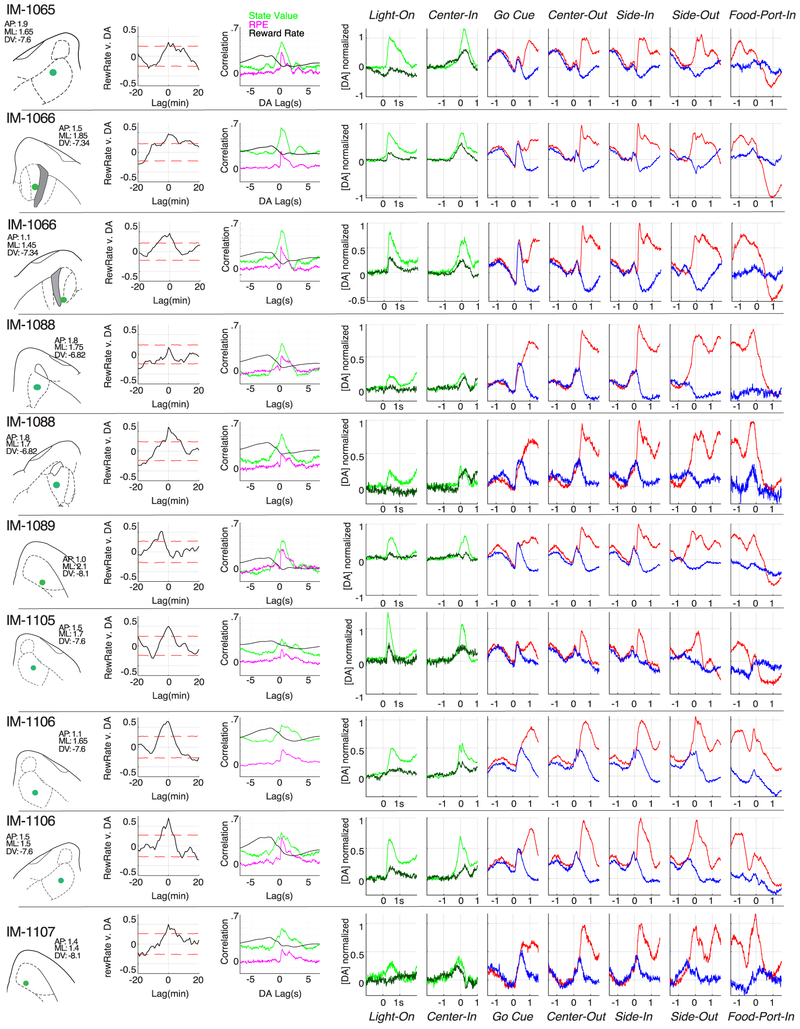
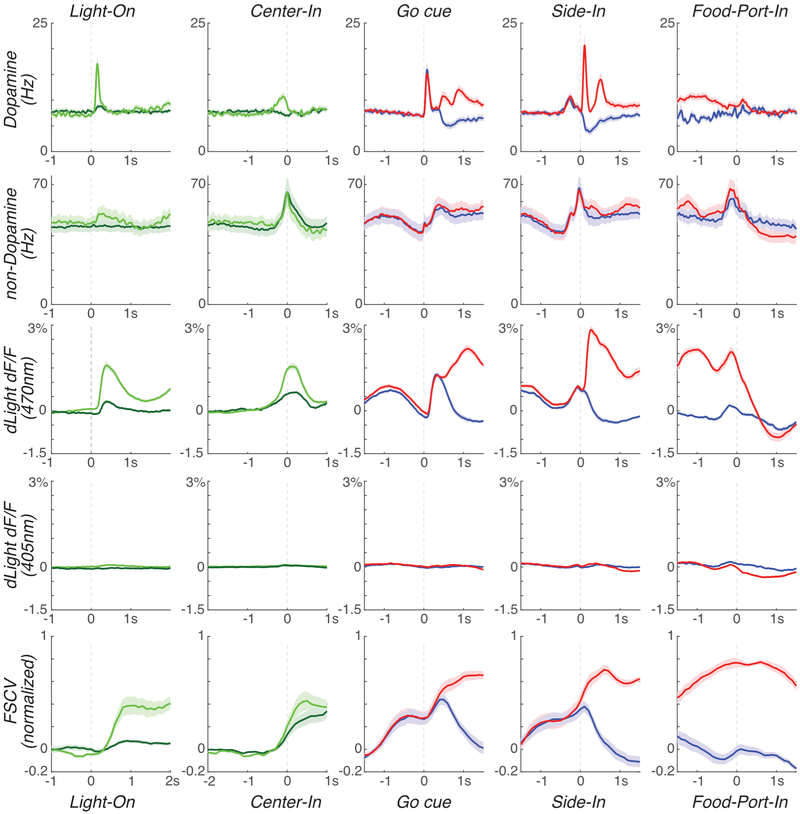
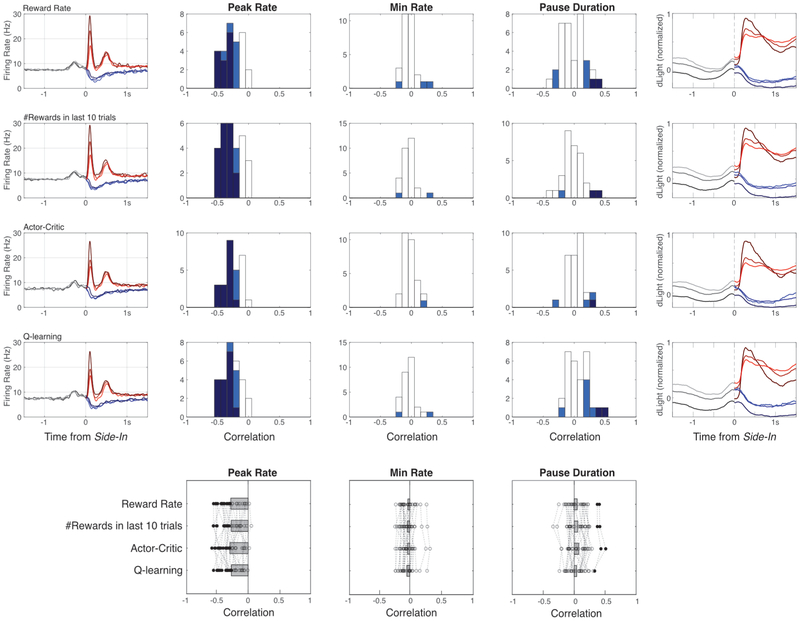
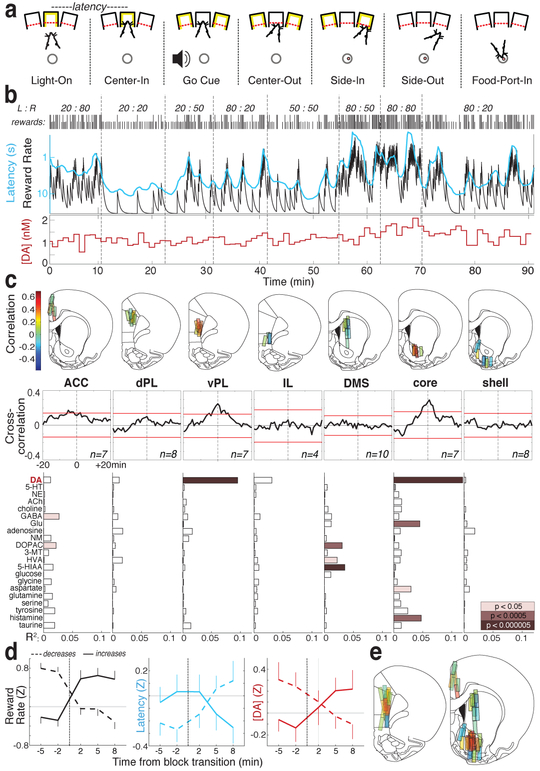
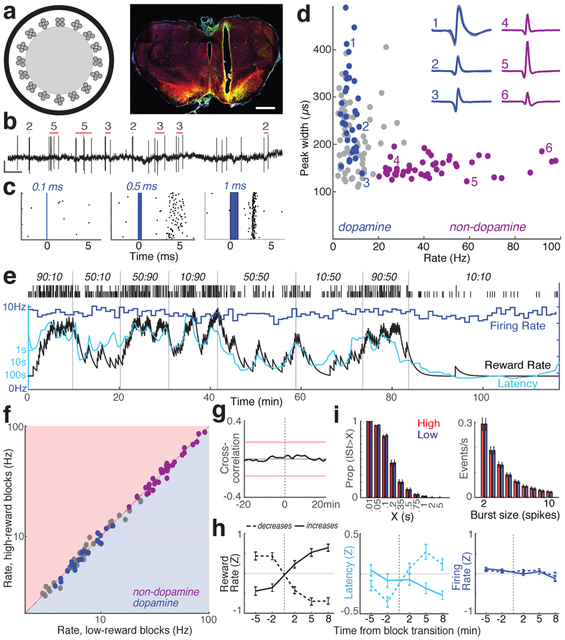
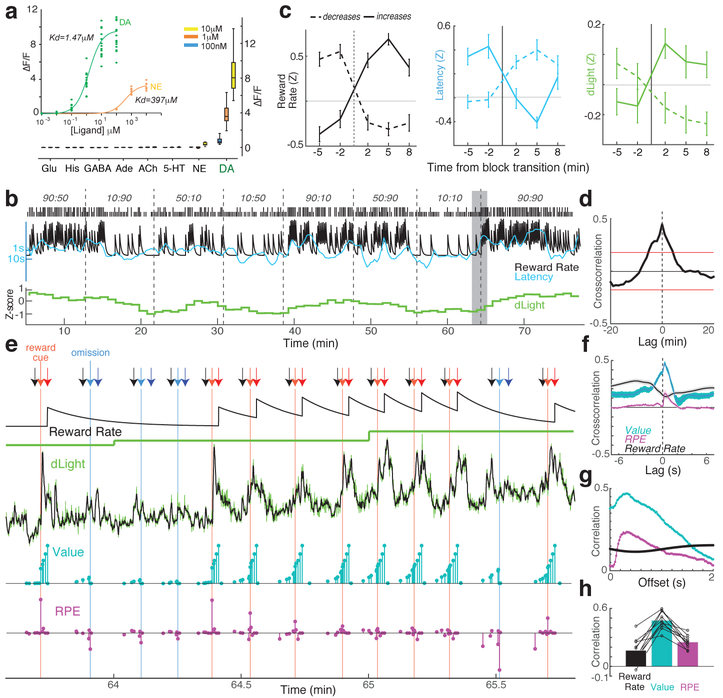
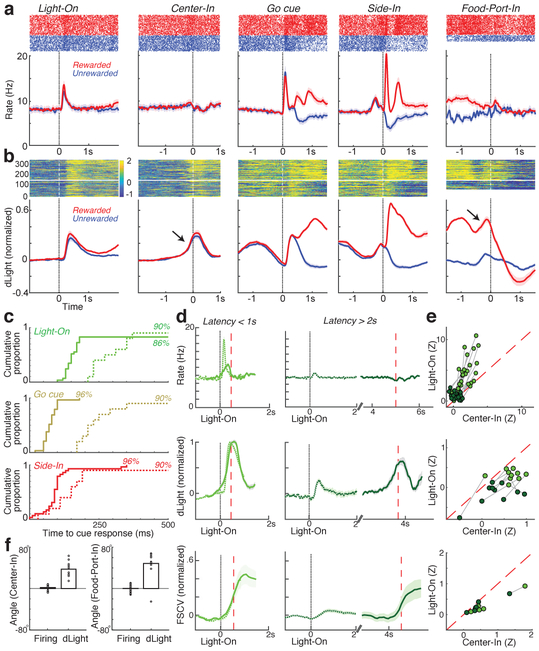
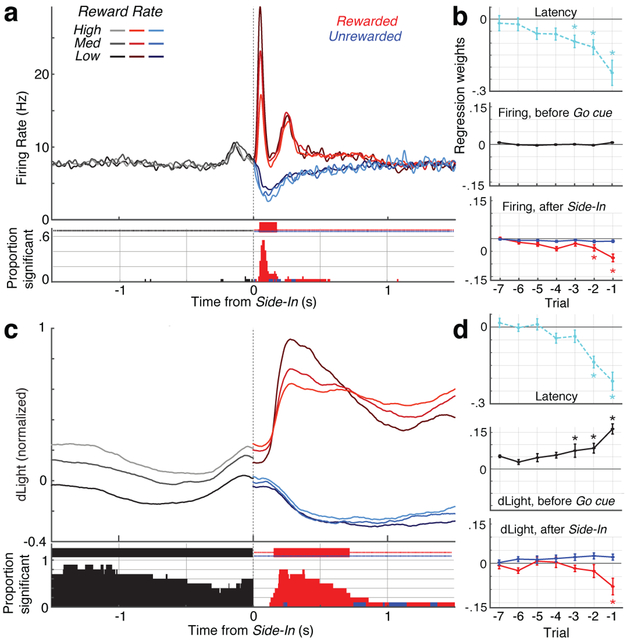
The dopamine projection from ventral tegmental area (VTA) to nucleus accumbens (NAc) is critical for motivation to work for rewards and reward-driven learning. How dopamine supports both functions is unclear. Dopamine cell spiking can encode prediction errors, which are vital learning signals in computational theories of adaptive behaviour. By contrast, dopamine release ramps up as animals approach rewards, mirroring reward expectation. This mismatch might reflect differences in behavioural tasks, slower changes in dopamine cell spiking or spike-independent modulation of dopamine release. Here we compare spiking of identified VTA dopamine cells with NAc dopamine release in the same decision-making task. Cues that indicate an upcoming reward increased both spiking and release. However, NAc core dopamine release also covaried with dynamically evolving reward expectations, without corresponding changes in VTA dopamine cell spiking. Our results suggest a fundamental difference in how dopamine release is regulated to achieve distinct functions: broadcast burst signals promote learning, whereas local control drives motivation.
Figures
Comment in
-
Closing in on what motivates motivation.Nature. 2019 Jun;570(7759):40-42. doi: 10.1038/d41586-019-01589-6. Nature. 2019. PMID: 31160742 No abstract available.
