

A tension-independent mechanism reduces Aurora B-mediated phosphorylation upon microtubule capture by CENP-E at the kinetochore
- PMID: 31122175
- PMCID: PMC6592239
- DOI: 10.1080/15384101.2019.1617615
A tension-independent mechanism reduces Aurora B-mediated phosphorylation upon microtubule capture by CENP-E at the kinetochore
Abstract
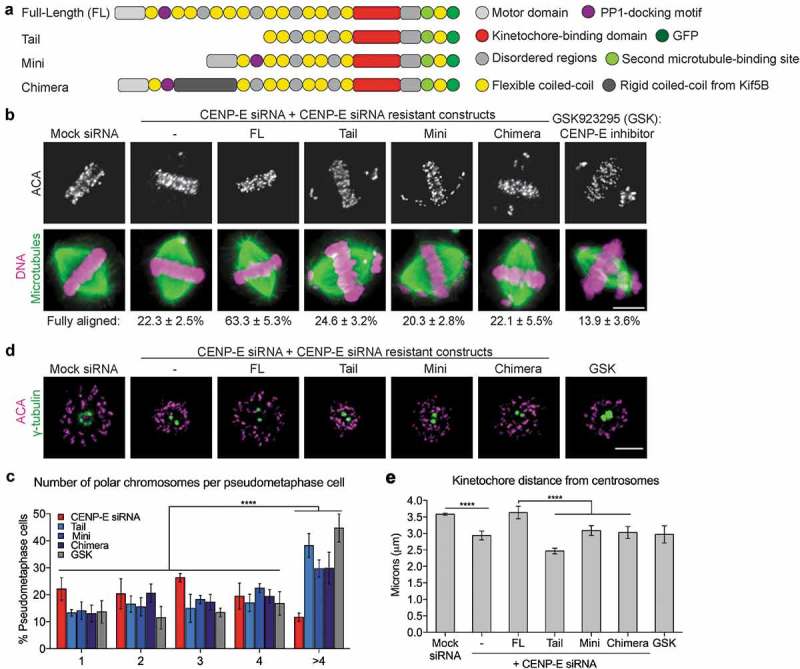
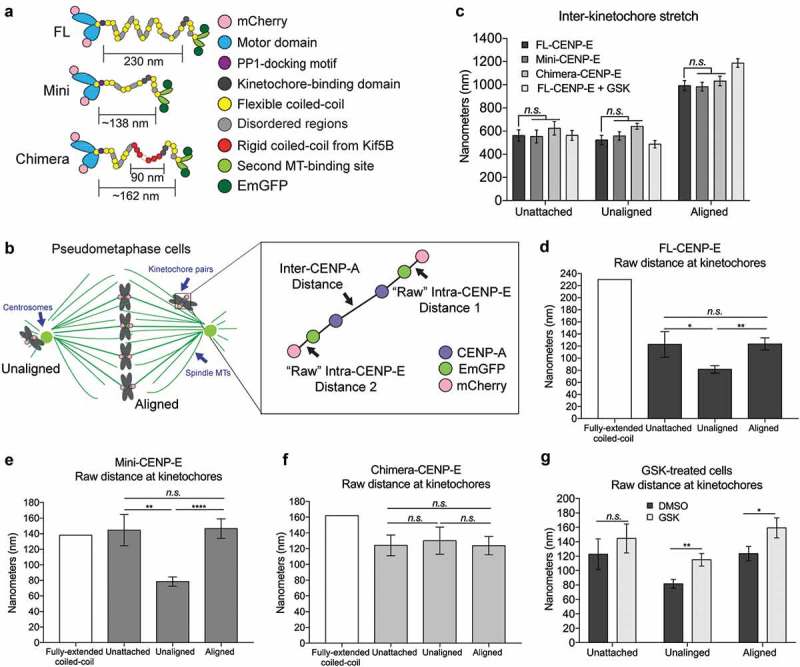
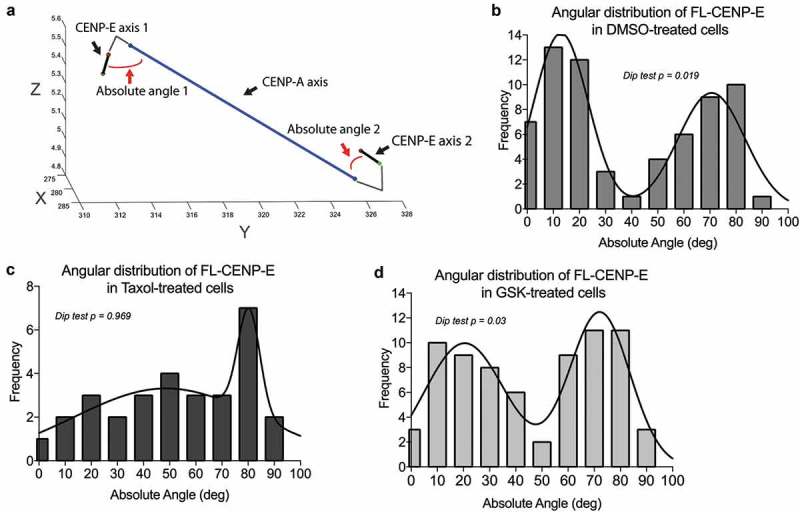
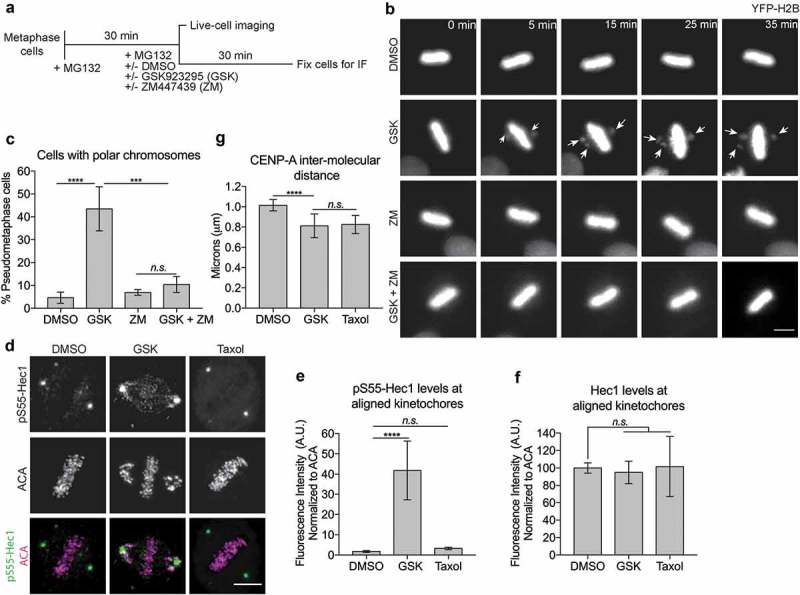
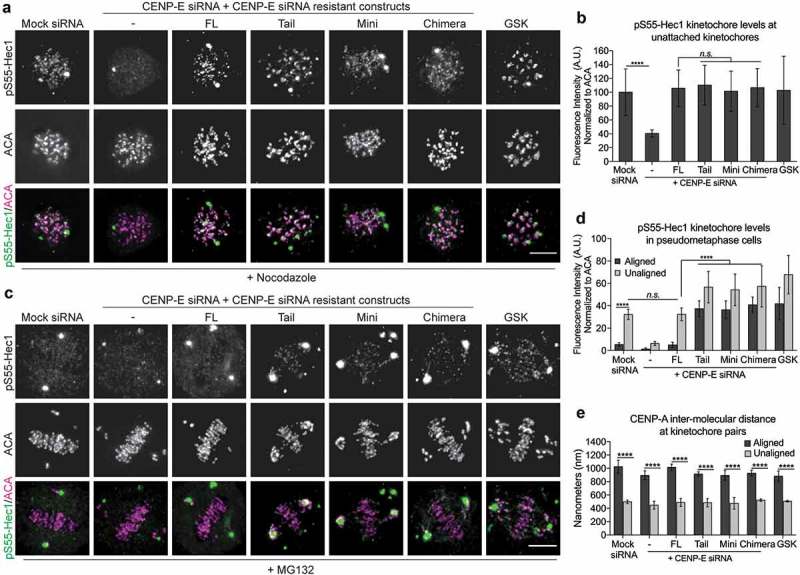
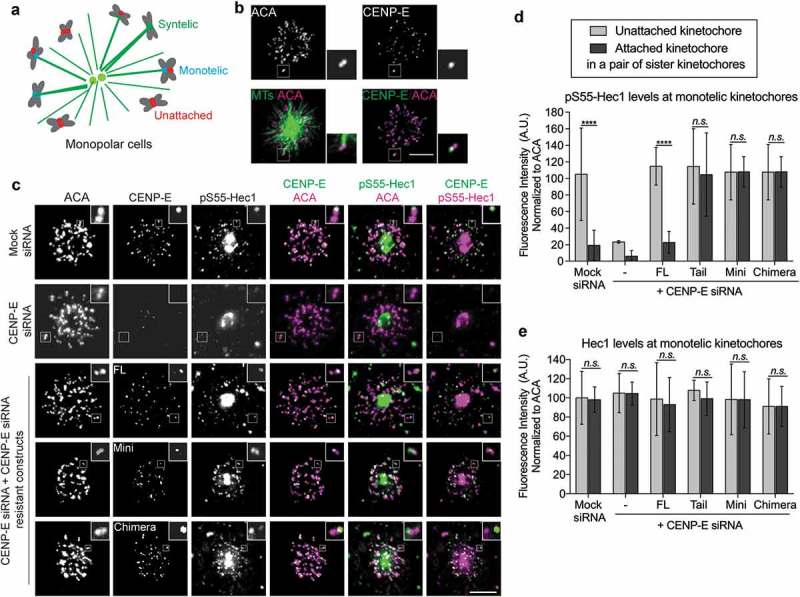
During mitosis, Aurora B kinase is required for forming proper bi-oriented kinetochore-microtubule attachments. Current models suggest that tension exerted between a pair of sister-kinetochores (inter-kinetochore stretch) produces a spatial separation of Aurora B kinase from kinetochore-associated microtubule binding substrates, such as the Knl1-Mis12-Ndc80 (KMN) network, resulting in a decrease of phosphorylation and, thus, an increase of affinity for microtubules. Using Single-Molecule High-Resolution Colocalization (SHREC) microscopy analysis of the kinetochore-associated motor CENP-E, we now show that CENP-E undergoes structural rearrangements prior to and after tension generation at the kinetochore, and displays a bi-modal Gaussian distribution on a pair of bi-oriented sister kinetochores. The conformational change of CENP-E depends on its microtubule-stimulated motor motility and the highly flexible coiled-coil between its motor and kinetochore-binding tail domains. Chemical inhibition of the motor motility or perturbations of the coiled-coil domain of CENP-E increases Aurora B-mediated Ndc80 phosphorylation in a tension-independent manner. Metaphase chromosome misalignment caused by CENP-E inhibition can be rescued by chemical inhibition of Aurora B kinase. Furthermore, a pair of monotelic sister-kinetochores shows asymmetric levels of Aurora B-mediated phosphorylation in mono-polar spindles depending on CENP-E motor activity. These results collectively suggest a tension-independent mechanism to reduce Aurora B-mediated phosphorylation of outer kinetochore components in response to microtubule capture by CENP-E.
Keywords: Aurora B; CENP-E; chromosome; kinetochore; microtubule; mitosis.
Figures
Similar articles
-
Phosphorylation of HsMis13 by Aurora B kinase is essential for assembly of functional kinetochore.J Biol Chem. 2008 Sep 26;283(39):26726-36. doi: 10.1074/jbc.M804207200. Epub 2008 Jul 17. J Biol Chem. 2008. PMID: 18640974 Free PMC article.
-
Mechanisms of kinesin-7 CENP-E in kinetochore-microtubule capture and chromosome alignment during cell division.Biol Cell. 2019 Jun;111(6):143-160. doi: 10.1111/boc.201800082. Epub 2019 Feb 26. Biol Cell. 2019. PMID: 30784092 Review.
-
Phosphorylation of CENP-C by Aurora B facilitates kinetochore attachment error correction in mitosis.Proc Natl Acad Sci U S A. 2017 Dec 12;114(50):E10667-E10676. doi: 10.1073/pnas.1710506114. Epub 2017 Nov 27. Proc Natl Acad Sci U S A. 2017. PMID: 29180432 Free PMC article.
-
Aurora A kinase phosphorylates Hec1 to regulate metaphase kinetochore-microtubule dynamics.J Cell Biol. 2018 Jan 2;217(1):163-177. doi: 10.1083/jcb.201707160. Epub 2017 Nov 29. J Cell Biol. 2018. PMID: 29187526 Free PMC article.
-
Correcting aberrant kinetochore microtubule attachments: a hidden regulation of Aurora B on microtubules.Curr Opin Cell Biol. 2019 Jun;58:34-41. doi: 10.1016/j.ceb.2018.12.007. Epub 2019 Jan 23. Curr Opin Cell Biol. 2019. PMID: 30684807 Free PMC article. Review.
Cited by
-
Kinesin-7 CENP-E regulates the formation and structural maintenance of the acrosome.Cell Tissue Res. 2021 Mar;383(3):1167-1182. doi: 10.1007/s00441-020-03341-3. Epub 2020 Nov 25. Cell Tissue Res. 2021. PMID: 33237480
-
The Mitotic Apparatus and Kinetochores in Microcephaly and Neurodevelopmental Diseases.Cells. 2019 Dec 24;9(1):49. doi: 10.3390/cells9010049. Cells. 2019. PMID: 31878213 Free PMC article. Review.
-
Leaving no-one behind: how CENP-E facilitates chromosome alignment.Essays Biochem. 2020 Sep 4;64(2):313-324. doi: 10.1042/EBC20190073. Essays Biochem. 2020. PMID: 32347304 Free PMC article. Review.
-
Kinesin-7 CENP-E mediates chromosome alignment and spindle assembly checkpoint in meiosis I.Chromosoma. 2024 Apr;133(2):149-168. doi: 10.1007/s00412-024-00818-w. Epub 2024 Mar 8. Chromosoma. 2024. PMID: 38456964
-
Kinesin-7 CENP-E in tumorigenesis: Chromosome instability, spindle assembly checkpoint, and applications.Front Mol Biosci. 2024 Mar 15;11:1366113. doi: 10.3389/fmolb.2024.1366113. eCollection 2024. Front Mol Biosci. 2024. PMID: 38560520 Free PMC article. Review.
References
-
- Walczak CE, Heald R. Mechanisms of mitotic spindle assembly and function. Int Rev Cytol. 2008;265:111–158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials