

Overexpression of Solanum habrochaites microRNA319d (sha-miR319d) confers chilling and heat stress tolerance in tomato (S. lycopersicum)
- PMID: 31122194
- PMCID: PMC6533698
- DOI: 10.1186/s12870-019-1823-x
Overexpression of Solanum habrochaites microRNA319d (sha-miR319d) confers chilling and heat stress tolerance in tomato (S. lycopersicum)
Abstract
Background: MicroRNA319 (miR319) acts as an essential regulator of gene expression during plant development and under stress conditions. Although the role of miR319a in regulating leaf development has been well studied in tomato (Solanum lycopersicum), the function of the recently discovered wild tomato Solanum habrochaites miRNA319d (sha-miR319d) remains poorly understood. In this study, we overexpressed sha-miR319d in cultivated tomato 'Micro-Tom' to further investigate its role in tomato temperature stress responses.
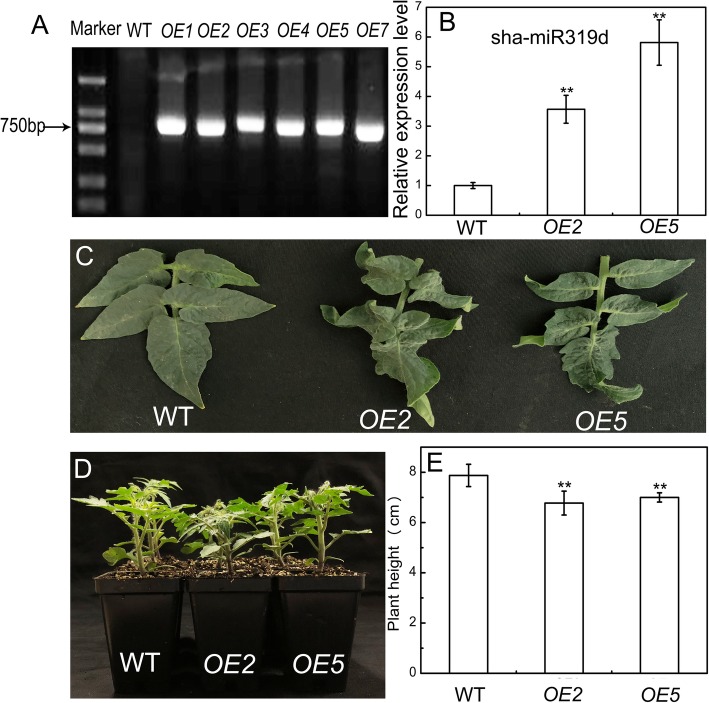
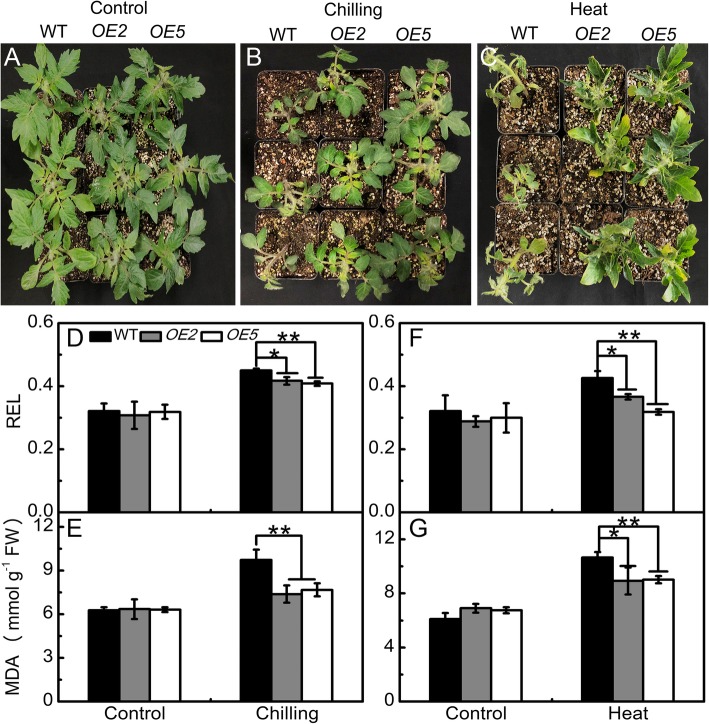
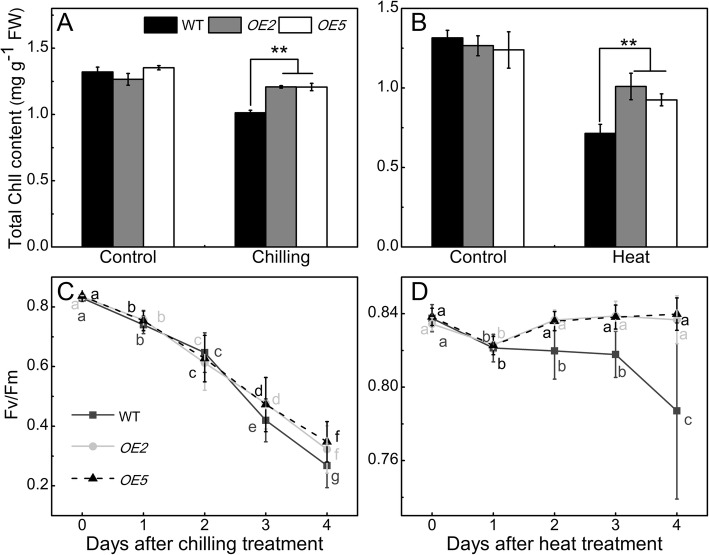
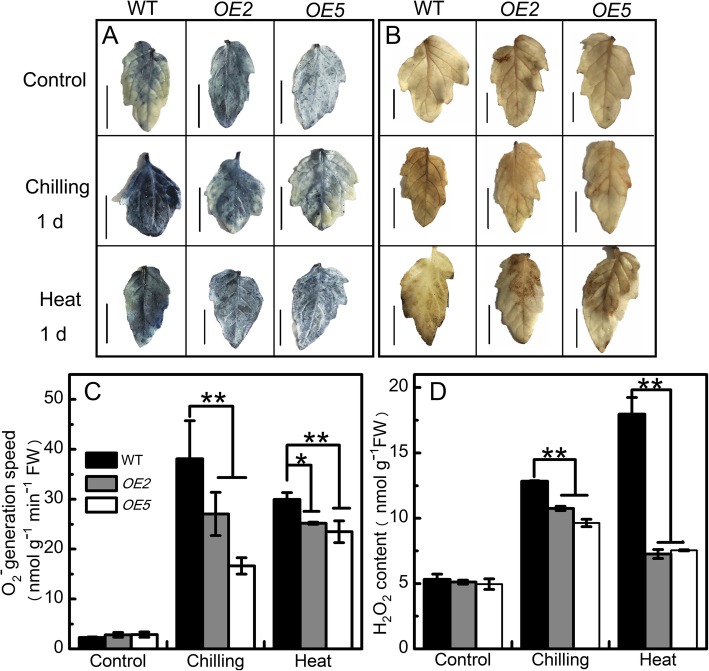
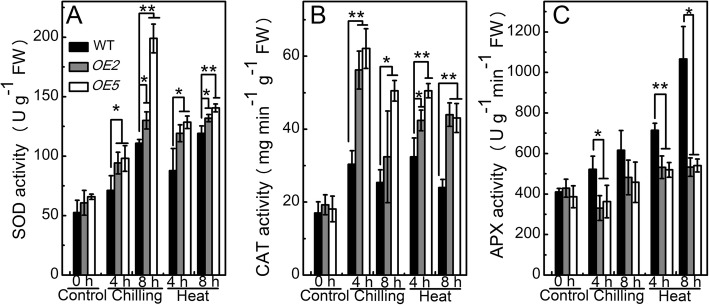
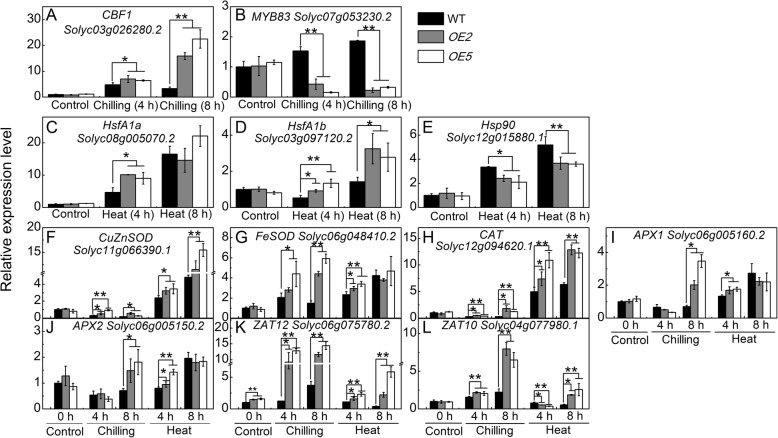
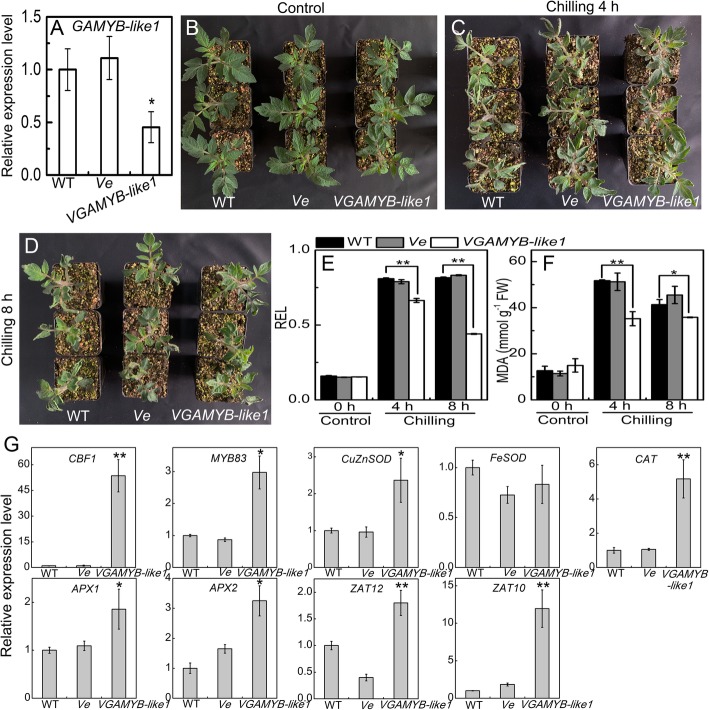
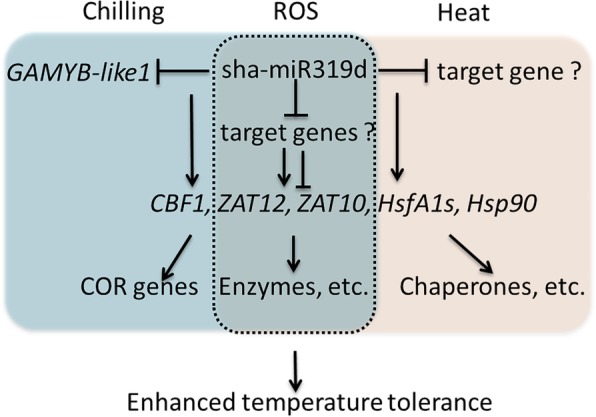
Results: Under chilling or heat stress, sha-miR319d-overexpressing plants showed enhanced stress tolerance, including lower relative electrolyte leakage (REL), malondialdehyde (MDA) concentration, O2- generation and H2O2 concentration and higher chlorophyll contents and Fv/Fm values than wild-type (WT) plants. Overexpression of sha-miR319d enhanced the activities of superoxide dismutase (SOD) and catalase (CAT), with possible correlation with elevated expression levels of the genes FeSOD, CuZnSOD and CAT. Moreover, different expression levels of key genes involved in chilling (MYB83 and CBF1), heat (HsfA1a, HsfA1b and Hsp90), and reactive oxygen species (ROS) (ZAT12 and ZAT10) signaling in transgenic plants and WT were determined, suggesting a role for sha-miR319d in regulating tomato temperature stress via chilling, heat and ROS signaling. Silencing GAMYB-like1 increased tomato chilling tolerance as well as the expression levels of CBF1, CuZnSOD, CAT, APX1, APX2, ZAT12 and ZAT10. Additionally, overexpression of sha-miR319d in tomato caused plant leaf crinkling and reduced height.
Conclusions: Overexpression of sha-miR319d confers chilling and heat stress tolerance in tomato. Sha-miR319d regulates tomato chilling tolerance, possibly by inhibiting expression of GAMYB-like1 and further alters chilling, heat and ROS signal transduction. Our research provides insight for further study of the role of sha-miR319d in tomato growth and stress regulation and lays a foundation for the genetic improvement of tomato.
Keywords: Chilling stress; Heat stress; Solanum habrochaites; Tomato; miR319.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Overexpression of chloroplast-localized small molecular heat-shock protein enhances chilling tolerance in tomato plant.Zhi Wu Sheng Li Yu Fen Zi Sheng Wu Xue Xue Bao. 2005 Apr;31(2):167-74. Zhi Wu Sheng Li Yu Fen Zi Sheng Wu Xue Xue Bao. 2005. PMID: 15840935
-
Enhanced brassinosteroid signaling via the overexpression of SlBRI1 positively regulates the chilling stress tolerance of tomato.Plant Sci. 2022 Jul;320:111281. doi: 10.1016/j.plantsci.2022.111281. Epub 2022 Apr 4. Plant Sci. 2022. PMID: 35643607
-
A comparison of the low temperature transcriptomes of two tomato genotypes that differ in freezing tolerance: Solanum lycopersicum and Solanum habrochaites.BMC Plant Biol. 2015 Jun 6;15:132. doi: 10.1186/s12870-015-0521-6. BMC Plant Biol. 2015. PMID: 26048292 Free PMC article.
-
MicroRNAs in tomato plants.Sci China Life Sci. 2011 Jul;54(7):599-605. doi: 10.1007/s11427-011-4188-4. Epub 2011 Jul 12. Sci China Life Sci. 2011. PMID: 21748583 Review.
-
The inheritance of chilling tolerance in tomato (Lycopersicon spp.).Plant Biol (Stuttg). 2005 Mar;7(2):118-30. doi: 10.1055/s-2005-837495. Plant Biol (Stuttg). 2005. PMID: 15822007 Review.
Cited by
-
Silencing of Sly-miR171d increased the expression of GRAS24 and enhanced postharvest chilling tolerance of tomato fruit.Front Plant Sci. 2022 Sep 9;13:1006940. doi: 10.3389/fpls.2022.1006940. eCollection 2022. Front Plant Sci. 2022. PMID: 36161008 Free PMC article.
-
Characterization of PsmiR319 during flower development in early- and late-flowering tree peonies cultivars.Plant Signal Behav. 2022 Dec 31;17(1):2120303. doi: 10.1080/15592324.2022.2120303. Plant Signal Behav. 2022. PMID: 36200538 Free PMC article.
-
Overexpression of Mtr-miR319a Contributes to Leaf Curl and Salt Stress Adaptation in Arabidopsis thaliana and Medicago truncatula.Int J Mol Sci. 2022 Dec 27;24(1):429. doi: 10.3390/ijms24010429. Int J Mol Sci. 2022. PMID: 36613873 Free PMC article.
-
Comparative Transcriptome Profiling Provides Insights into Plant Salt Tolerance in Watermelon (Citrullus lanatus).Life (Basel). 2022 Jul 12;12(7):1033. doi: 10.3390/life12071033. Life (Basel). 2022. PMID: 35888121 Free PMC article.
-
SlIAA9 Mutation Maintains Photosynthetic Capabilities under Heat-Stress Conditions.Plants (Basel). 2023 Jan 13;12(2):378. doi: 10.3390/plants12020378. Plants (Basel). 2023. PMID: 36679090 Free PMC article.
References
-
- Huang SC, Lu GH, Tang CY, Ji YJ, Tan GS, Hu DQ, Cheng J, Wang GH, Qi JL, Yang YH. Identification and comparative analysis of aluminum-induced microRNAs conferring plant tolerance to aluminum stress in soybean. Biol Plant. 2017:1–12.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
