

Comprehensive Multiple eQTL Detection and Its Application to GWAS Interpretation
- PMID: 31123039
- PMCID: PMC6614888
- DOI: 10.1534/genetics.119.302091
Comprehensive Multiple eQTL Detection and Its Application to GWAS Interpretation
Abstract
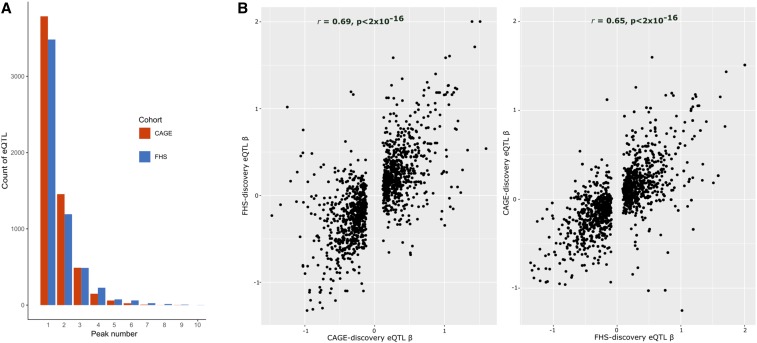
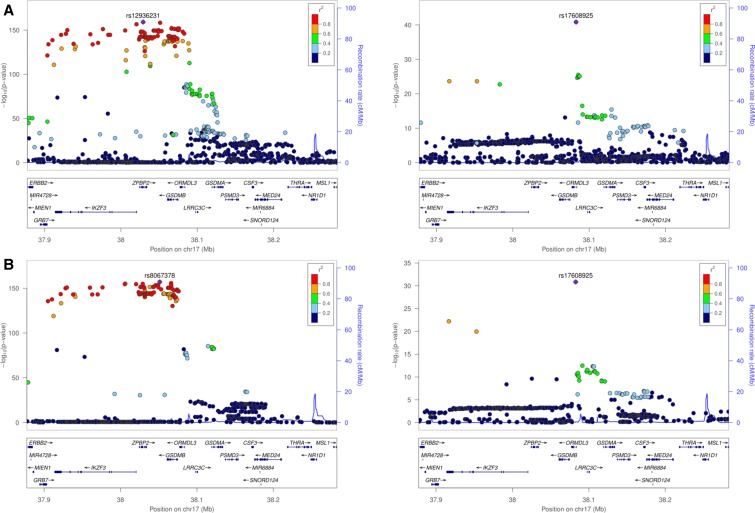
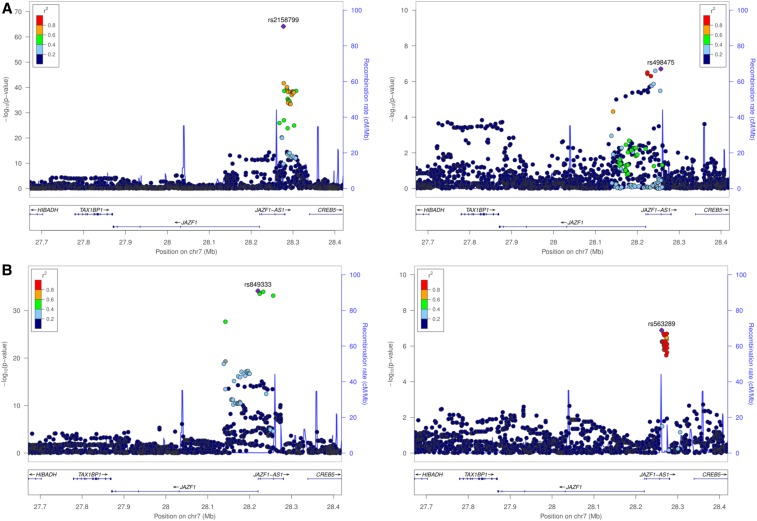
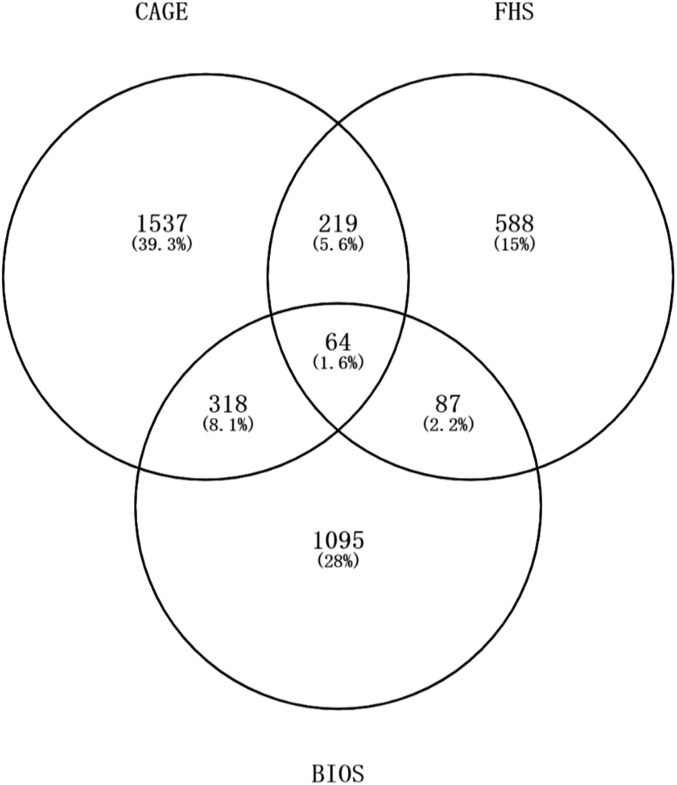
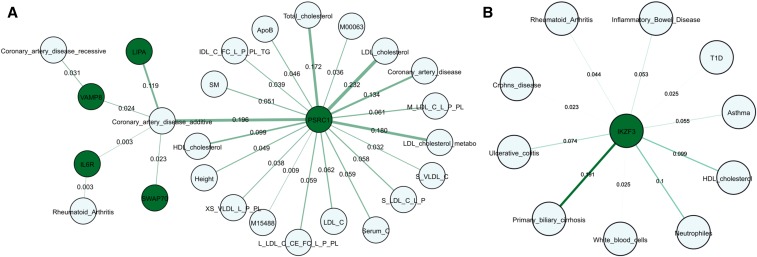
Expression QTL (eQTL) detection has emerged as an important tool for unraveling the relationship between genetic risk factors and disease or clinical phenotypes. Most studies are predicated on the assumption that only a single causal variant explains the association signal in each interval. This greatly simplifies the statistical modeling, but is liable to biases in scenarios where multiple local causal-variants are responsible. Here, our primary goal was to address the prevalence of secondary cis-eQTL signals regulating peripheral blood gene expression locally, utilizing two large human cohort studies, each >2500 samples with accompanying whole genome genotypes. The CAGE (Consortium for the Architecture of Gene Expression) dataset is a compendium of Illumina microarray studies, and the Framingham Heart Study is a two-generation Affymetrix dataset. We also describe Bayesian colocalization analysis of the extent of sharing of cis-eQTL detected in both studies as well as with the BIOS RNAseq dataset. Stepwise conditional modeling demonstrates that multiple eQTL signals are present for ∼40% of over 3500 eGenes in both microarray datasets, and that the number of loci with additional signals reduces by approximately two-thirds with each conditioning step. Although <20% of the peak signals across platforms fine map to the same credible interval, the colocalization analysis finds that as many as 50-60% of the primary eQTL are actually shared. Subsequently, colocalization of eQTL signals with GWAS hits detected 1349 genes whose expression in peripheral blood is associated with 591 human phenotype traits or diseases, including enrichment for genes with regulatory functions. At least 10%, and possibly as many as 40%, of eQTL-trait colocalized signals are due to nonprimary cis-eQTL peaks, but just one-quarter of these colocalization signals replicated across the gene expression datasets. Our results are provided as a web-based resource for visualization of multi-site regulation of gene expression and its association with human complex traits and disease states.
Keywords: PolyQTL; colocalization; conditional association; fine mapping; gene regulation; linkage disequilibrium.
Copyright © 2019 by the Genetics Society of America.
Figures

Similar articles
-
Colocalization of GWAS and eQTL signals at loci with multiple signals identifies additional candidate genes for body fat distribution.Hum Mol Genet. 2019 Dec 15;28(24):4161-4172. doi: 10.1093/hmg/ddz263. Hum Mol Genet. 2019. PMID: 31691812 Free PMC article.
-
A mega-analysis of expression quantitative trait loci in retinal tissue.PLoS Genet. 2020 Sep 1;16(9):e1008934. doi: 10.1371/journal.pgen.1008934. eCollection 2020 Sep. PLoS Genet. 2020. PMID: 32870927 Free PMC article.
-
Genetic control of gene expression at novel and established chronic obstructive pulmonary disease loci.Hum Mol Genet. 2015 Feb 15;24(4):1200-10. doi: 10.1093/hmg/ddu525. Epub 2014 Oct 14. Hum Mol Genet. 2015. PMID: 25315895 Free PMC article.
-
Towards the Genetic Architecture of Complex Gene Expression Traits: Challenges and Prospects for eQTL Mapping in Humans.Genes (Basel). 2022 Jan 26;13(2):235. doi: 10.3390/genes13020235. Genes (Basel). 2022. PMID: 35205280 Free PMC article. Review.
-
Expression QTLs Mapping and Analysis: A Bayesian Perspective.Methods Mol Biol. 2017;1488:189-215. doi: 10.1007/978-1-4939-6427-7_8. Methods Mol Biol. 2017. PMID: 27933525 Review.
Cited by
-
Genotype inference from aggregated chromatin accessibility data reveals genetic regulatory mechanisms.bioRxiv [Preprint]. 2024 Sep 5:2024.09.04.610850. doi: 10.1101/2024.09.04.610850. bioRxiv. 2024. Update in: Genome Biol. 2025 Mar 30;26(1):81. doi: 10.1186/s13059-025-03538-1. PMID: 39282458 Free PMC article. Updated. Preprint.
-
The Genetic Control of the Rheumatic Heart: Closing the Genotype-Phenotype Gap.Front Med (Lausanne). 2021 Mar 24;8:611036. doi: 10.3389/fmed.2021.611036. eCollection 2021. Front Med (Lausanne). 2021. PMID: 33842495 Free PMC article. Review.
-
Where Are the Disease-Associated eQTLs?Trends Genet. 2021 Feb;37(2):109-124. doi: 10.1016/j.tig.2020.08.009. Epub 2020 Sep 7. Trends Genet. 2021. PMID: 32912663 Free PMC article. Review.
-
Fine-mapping within eQTL credible intervals by expression CROP-seq.Biol Methods Protoc. 2020 Mar 28;5(1):bpaa008. doi: 10.1093/biomethods/bpaa008. eCollection 2020. Biol Methods Protoc. 2020. PMID: 32665975 Free PMC article.
-
Regulatory SNPs: Altered Transcription Factor Binding Sites Implicated in Complex Traits and Diseases.Int J Mol Sci. 2021 Jun 16;22(12):6454. doi: 10.3390/ijms22126454. Int J Mol Sci. 2021. PMID: 34208629 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
