

Tolerance of DNA Replication Stress Is Promoted by Fumarate Through Modulation of Histone Demethylation and Enhancement of Replicative Intermediate Processing in Saccharomyces cerevisiae
- PMID: 31123043
- PMCID: PMC6614904
- DOI: 10.1534/genetics.119.302238
Tolerance of DNA Replication Stress Is Promoted by Fumarate Through Modulation of Histone Demethylation and Enhancement of Replicative Intermediate Processing in Saccharomyces cerevisiae
Abstract
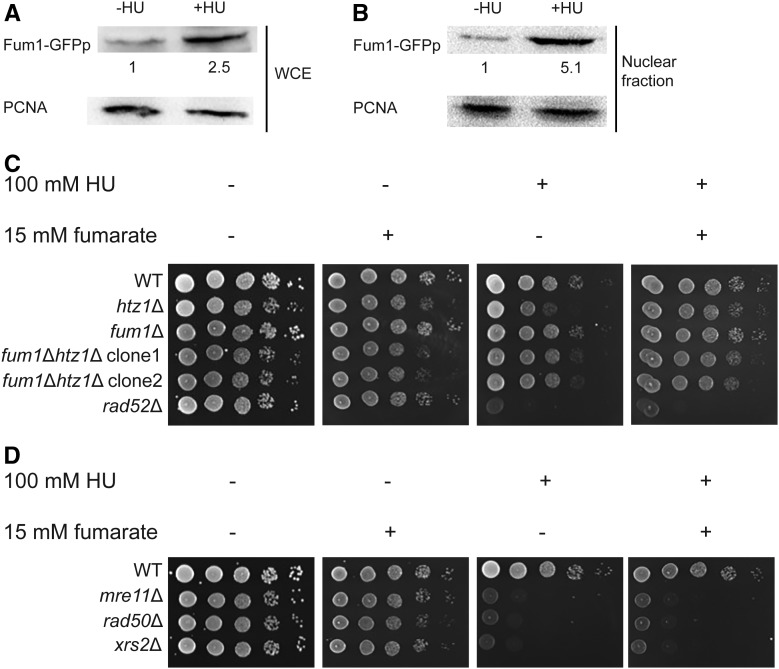
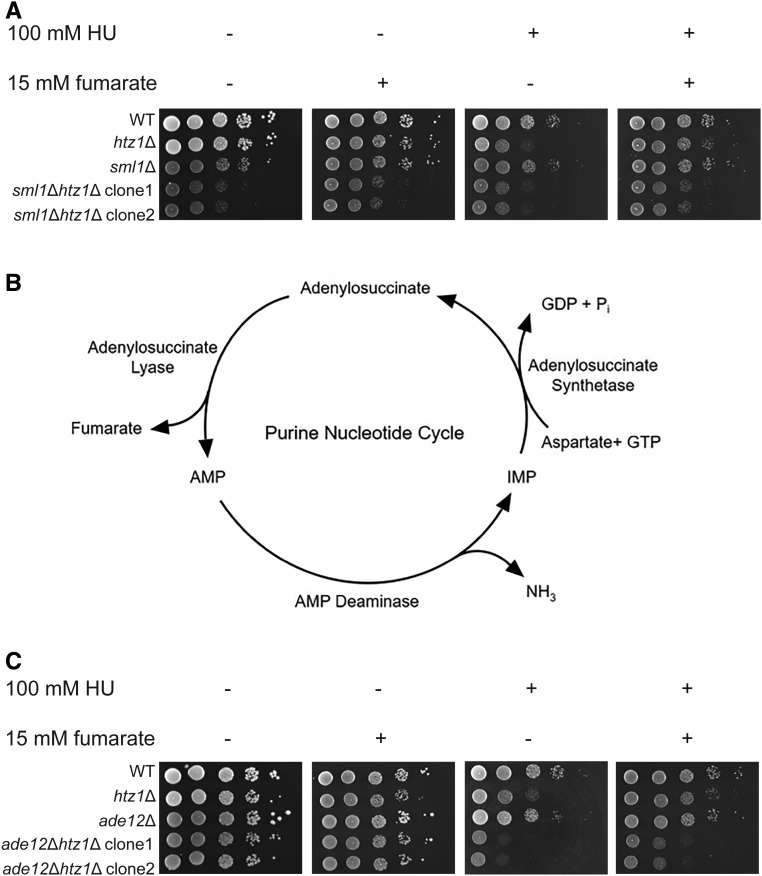
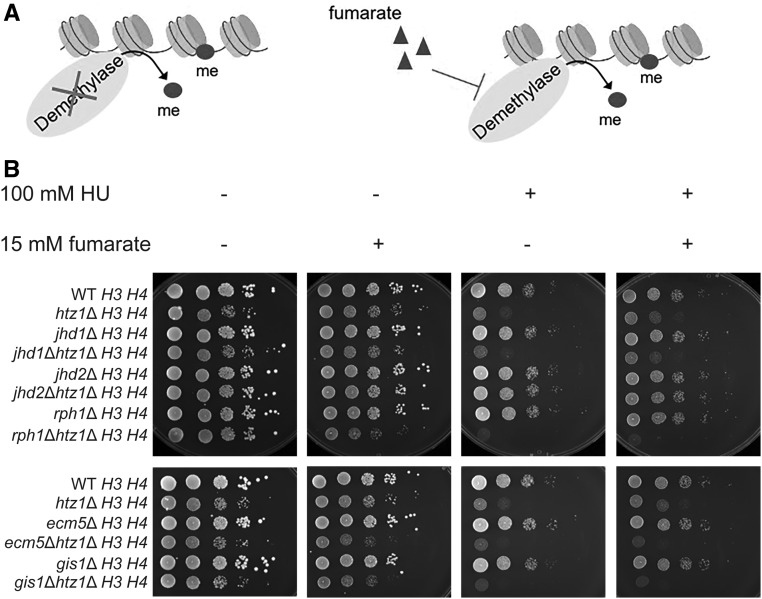
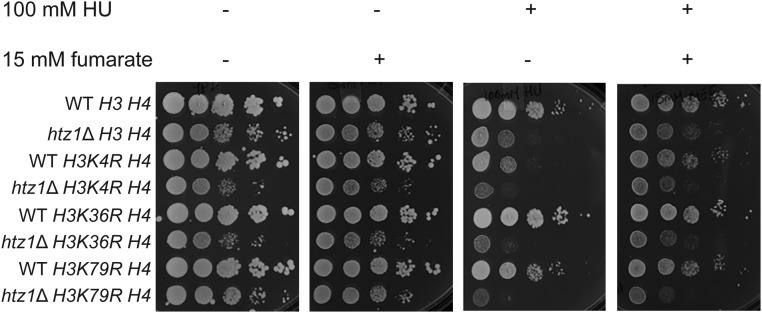
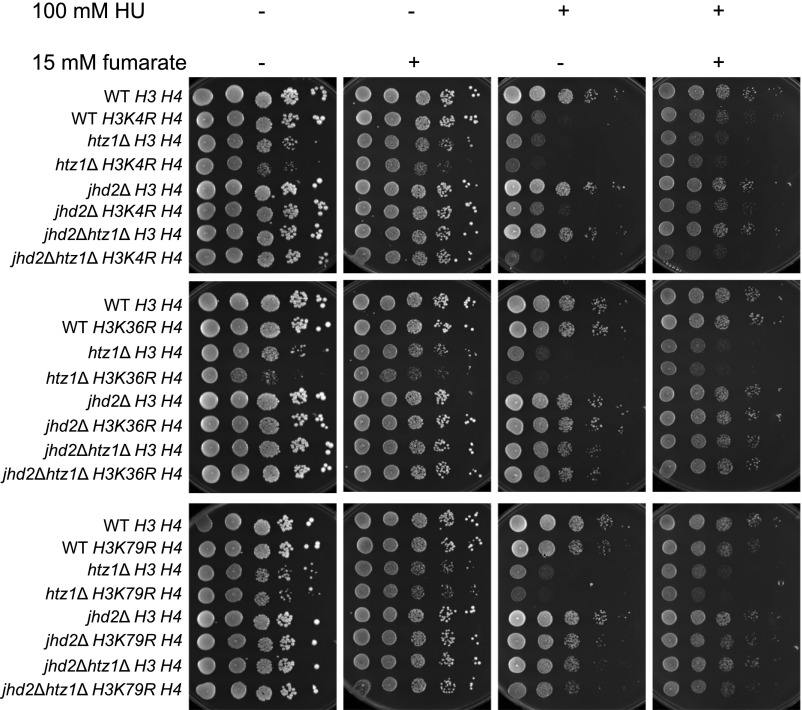
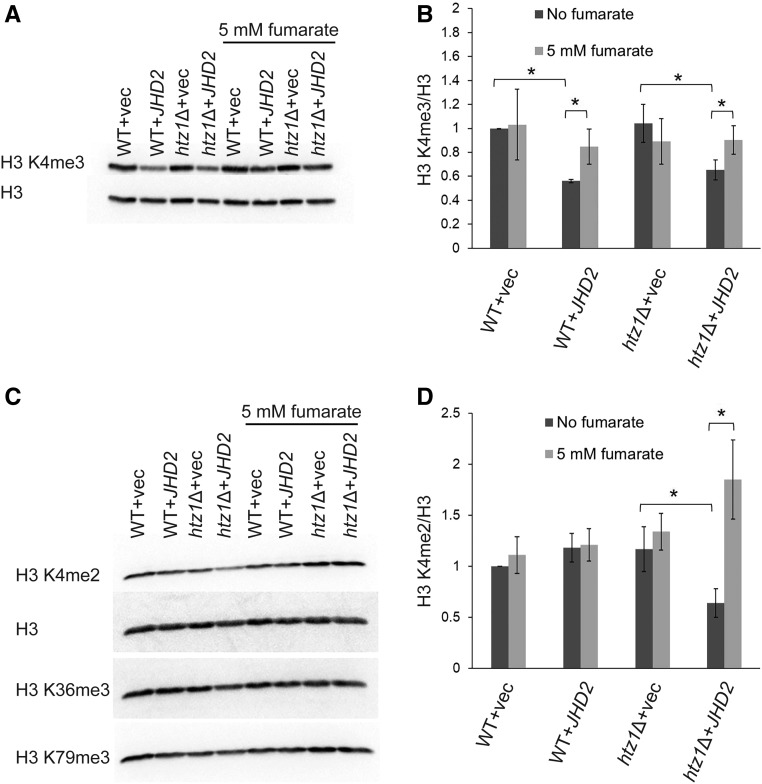
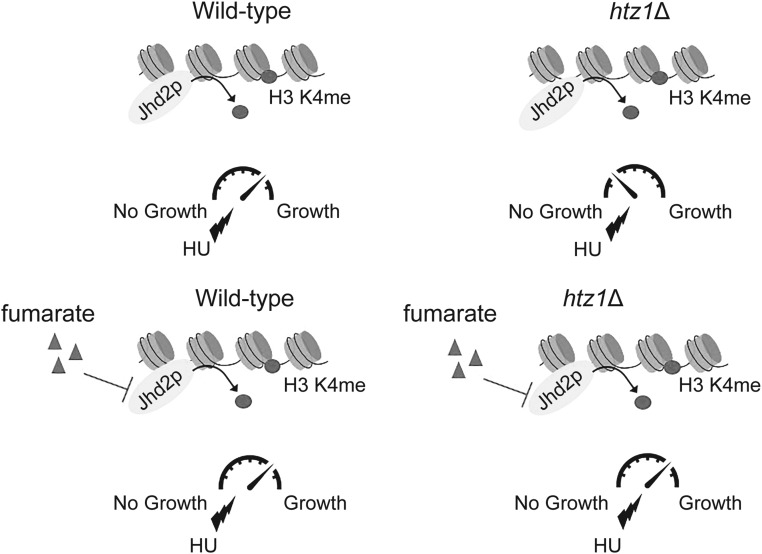
Fumarase is a well-characterized TCA cycle enzyme that catalyzes the reversible conversion of fumarate to malate. In mammals, fumarase acts as a tumor suppressor, and loss-of-function mutations in the FH gene in hereditary leiomyomatosis and renal cell cancer result in the accumulation of intracellular fumarate-an inhibitor of α-ketoglutarate-dependent dioxygenases. Fumarase promotes DNA repair by nonhomologous end joining in mammalian cells through interaction with the histone variant H2A.Z, and inhibition of KDM2B, a H3 K36-specific histone demethylase. Here, we report that Saccharomyces cerevisiae fumarase, Fum1p, acts as a response factor during DNA replication stress, and fumarate enhances survival of yeast lacking Htz1p (H2A.Z in mammals). We observed that exposure to DNA replication stress led to upregulation as well as nuclear enrichment of Fum1p, and raising levels of fumarate in cells via deletion of FUM1 or addition of exogenous fumarate suppressed the sensitivity to DNA replication stress of htz1Δ mutants. This suppression was independent of modulating nucleotide pool levels. Rather, our results are consistent with fumarate conferring resistance to DNA replication stress in htz1Δ mutants by inhibiting the H3 K4-specific histone demethylase Jhd2p, and increasing H3 K4 methylation. Although the timing of checkpoint activation and deactivation remained largely unaffected by fumarate, sensors and mediators of the DNA replication checkpoint were required for fumarate-dependent resistance to replication stress in the htz1Δ mutants. Together, our findings imply metabolic enzymes and metabolites aid in processing replicative intermediates by affecting chromatin modification states, thereby promoting genome integrity.
Keywords: DNA replication stress; HTZ1; JHD2; fumarate; histone methylation.
Copyright © 2019 by the Genetics Society of America.
Figures



Similar articles
-
Local generation of fumarate promotes DNA repair through inhibition of histone H3 demethylation.Nat Cell Biol. 2015 Sep;17(9):1158-68. doi: 10.1038/ncb3209. Epub 2015 Aug 3. Nat Cell Biol. 2015. PMID: 26237645 Free PMC article.
-
Fumarase: a mitochondrial metabolic enzyme and a cytosolic/nuclear component of the DNA damage response.PLoS Biol. 2010 Mar 9;8(3):e1000328. doi: 10.1371/journal.pbio.1000328. PLoS Biol. 2010. PMID: 20231875 Free PMC article.
-
Interaction of the Jhd2 Histone H3 Lys-4 Demethylase with Chromatin Is Controlled by Histone H2A Surfaces and Restricted by H2B Ubiquitination.J Biol Chem. 2015 Nov 27;290(48):28760-77. doi: 10.1074/jbc.M115.693085. Epub 2015 Oct 8. J Biol Chem. 2015. PMID: 26451043 Free PMC article.
-
The role of metabolic enzymes in mesenchymal tumors and tumor syndromes: genetics, pathology, and molecular mechanisms.Lab Invest. 2018 Apr;98(4):414-426. doi: 10.1038/s41374-017-0003-6. Epub 2018 Jan 16. Lab Invest. 2018. PMID: 29339836 Review.
-
Krebs cycle dysfunction shapes epigenetic landscape of chromatin: novel insights into mitochondrial regulation of aging process.Cell Signal. 2014 Jul;26(7):1598-603. doi: 10.1016/j.cellsig.2014.03.030. Epub 2014 Apr 2. Cell Signal. 2014. PMID: 24704120 Review.
Cited by
-
Hereditary Leiomyomatosis and Renal Cell Cancer: Recent Insights Into Mechanisms and Systemic Treatment.Front Oncol. 2021 May 25;11:686556. doi: 10.3389/fonc.2021.686556. eCollection 2021. Front Oncol. 2021. PMID: 34113573 Free PMC article. Review.
-
Tricarboxylic Acid Cycle Relationships with Non-Metabolic Processes: A Short Story with DNA Repair and Its Consequences on Cancer Therapy Resistance.Int J Mol Sci. 2024 Aug 21;25(16):9054. doi: 10.3390/ijms25169054. Int J Mol Sci. 2024. PMID: 39201738 Free PMC article. Review.
-
Unconventional metabolites in chromatin regulation.Biosci Rep. 2022 Jan 28;42(1):BSR20211558. doi: 10.1042/BSR20211558. Biosci Rep. 2022. PMID: 34988581 Free PMC article.
-
Fumarase affects the deoxyribonucleic acid damage response by protecting the mitochondrial desulfurase Nfs1p from modification and inactivation.iScience. 2021 Oct 26;24(11):103354. doi: 10.1016/j.isci.2021.103354. eCollection 2021 Nov 19. iScience. 2021. PMID: 34805801 Free PMC article.
-
Mitochondrial Factors in the Cell Nucleus.Int J Mol Sci. 2023 Sep 4;24(17):13656. doi: 10.3390/ijms241713656. Int J Mol Sci. 2023. PMID: 37686461 Free PMC article. Review.
References
-
- Adams A., Gottschling D. E., Kaiser C., Stearns T., 1998. Methods in Yeast Genetics : A Cold Spring Harbor Laboratory Course Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
