

Default Activation and Nuclear Translocation of the Plant Cellular Energy Sensor SnRK1 Regulate Metabolic Stress Responses and Development
- PMID: 31123051
- PMCID: PMC6635846
- DOI: 10.1105/tpc.18.00500
Default Activation and Nuclear Translocation of the Plant Cellular Energy Sensor SnRK1 Regulate Metabolic Stress Responses and Development
Abstract
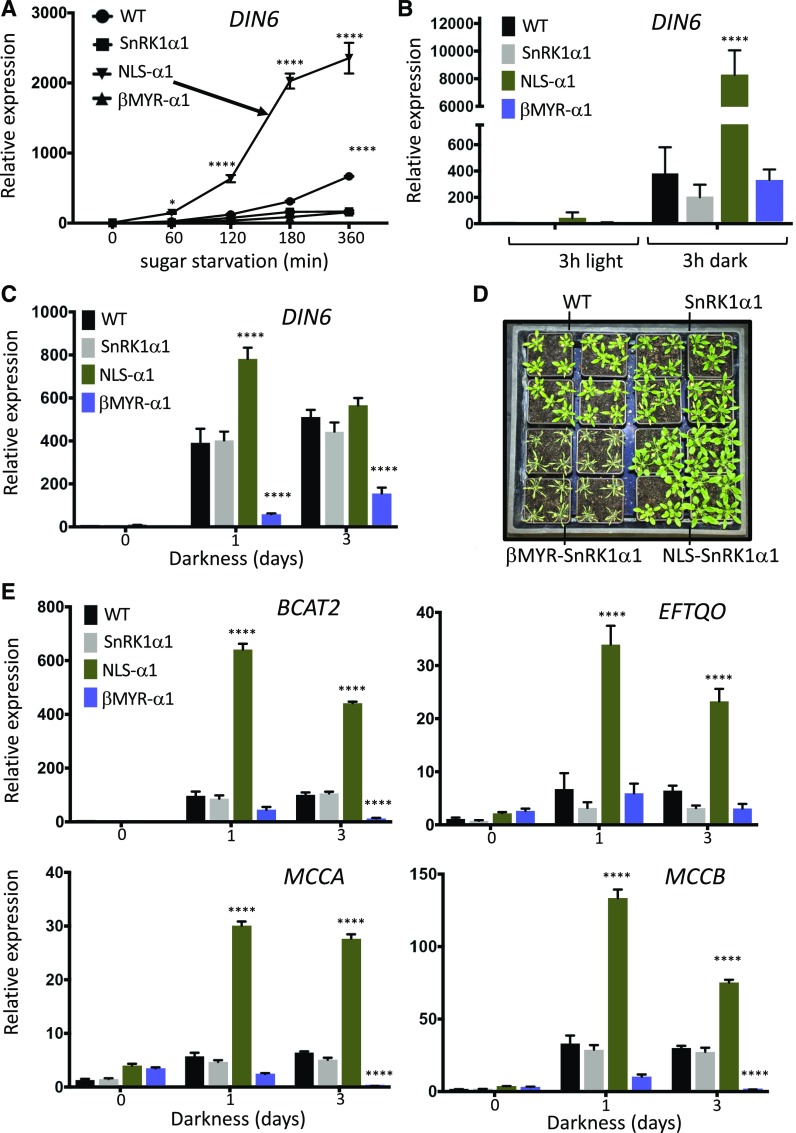
Energy homeostasis is vital to all living organisms. In eukaryotes, this process is controlled by fuel gauging protein kinases: AMP-activated kinase in mammals, Sucrose Non-Fermenting1 (SNF1) in yeast (Saccharomyces cerevisiae), and SNF1-related kinase1 (SnRK1) in plants. These kinases are highly conserved in structure and function and (according to this paradigm) operate as heterotrimeric complexes of catalytic-α and regulatory β- and γ-subunits, responding to low cellular nucleotide charge. Here, we determined that the Arabidopsis (Arabidopsis thaliana) SnRK1 catalytic α-subunit has regulatory subunit-independent activity, which is consistent with default activation (and thus controlled repression), a strategy more generally used by plants. Low energy stress (caused by darkness, inhibited photosynthesis, or hypoxia) also triggers SnRK1α nuclear translocation, thereby controlling induced but not repressed target gene expression to replenish cellular energy for plant survival. The myristoylated and membrane-associated regulatory β-subunits restrict nuclear localization and inhibit target gene induction. Transgenic plants with forced SnRK1α-subunit localization consistently were affected in metabolic stress responses, but their analysis also revealed key roles for nuclear SnRK1 in leaf and root growth and development. Our findings suggest that plants have modified the ancient, highly conserved eukaryotic energy sensor to better fit their unique lifestyle and to more effectively cope with changing environmental conditions.
© 2019 American Society of Plant Biologists. All rights reserved.
Figures








Similar articles
-
Dual and dynamic intracellular localization of Arabidopsis thaliana SnRK1.1.J Exp Bot. 2019 Apr 15;70(8):2325-2338. doi: 10.1093/jxb/erz023. J Exp Bot. 2019. PMID: 30753728
-
The hybrid four-CBS-domain KINβγ subunit functions as the canonical γ subunit of the plant energy sensor SnRK1.Plant J. 2013 Jul;75(1):11-25. doi: 10.1111/tpj.12192. Epub 2013 May 15. Plant J. 2013. PMID: 23551663 Free PMC article.
-
SnRK1 from Arabidopsis thaliana is an atypical AMPK.Plant J. 2015 Apr;82(2):183-92. doi: 10.1111/tpj.12813. Plant J. 2015. PMID: 25736509
-
Convergent energy and stress signaling.Trends Plant Sci. 2008 Sep;13(9):474-82. doi: 10.1016/j.tplants.2008.06.006. Epub 2008 Aug 11. Trends Plant Sci. 2008. PMID: 18701338 Free PMC article. Review.
-
The plant energy sensor: evolutionary conservation and divergence of SnRK1 structure, regulation, and function.J Exp Bot. 2016 Dec;67(22):6215-6252. doi: 10.1093/jxb/erw416. J Exp Bot. 2016. PMID: 27856705 Review.
Cited by
-
Low nitrogen conditions accelerate flowering by modulating the phosphorylation state of FLOWERING BHLH 4 in Arabidopsis.Proc Natl Acad Sci U S A. 2021 May 11;118(19):e2022942118. doi: 10.1073/pnas.2022942118. Proc Natl Acad Sci U S A. 2021. PMID: 33963081 Free PMC article.
-
Carbon/nitrogen metabolism and stress response networks - calcium-dependent protein kinases as the missing link?J Exp Bot. 2021 May 28;72(12):4190-4201. doi: 10.1093/jxb/erab136. J Exp Bot. 2021. PMID: 33787877 Free PMC article. Review.
-
Phosphorylation of S11 in PHR1 negatively controls its transcriptional activity.Physiol Plant. 2022 Nov;174(6):e13831. doi: 10.1111/ppl.13831. Physiol Plant. 2022. PMID: 36444477 Free PMC article.
-
Regulatory functions of cellular energy sensor SnRK1 for nitrate signalling through NLP7 repression.Nat Plants. 2022 Sep;8(9):1094-1107. doi: 10.1038/s41477-022-01236-5. Epub 2022 Sep 1. Nat Plants. 2022. PMID: 36050463
-
Convergence and Divergence of Sugar and Cytokinin Signaling in Plant Development.Int J Mol Sci. 2021 Jan 28;22(3):1282. doi: 10.3390/ijms22031282. Int J Mol Sci. 2021. PMID: 33525430 Free PMC article. Review.
References
-
- Baena-González E., Hanson J. (2017). Shaping plant development through the SnRK1-TOR metabolic regulators. Curr. Opin. Plant Biol. 35: 152–157. - PubMed
-
- Baena-González E., Rolland F., Thevelein J.M., Sheen J. (2007). A central integrator of transcription networks in plant stress and energy signalling. Nature 448: 938–942. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
