

One minute analysis of 200 histone posttranslational modifications by direct injection mass spectrometry
- PMID: 31123082
- PMCID: PMC6581051
- DOI: 10.1101/gr.247353.118
One minute analysis of 200 histone posttranslational modifications by direct injection mass spectrometry
Abstract
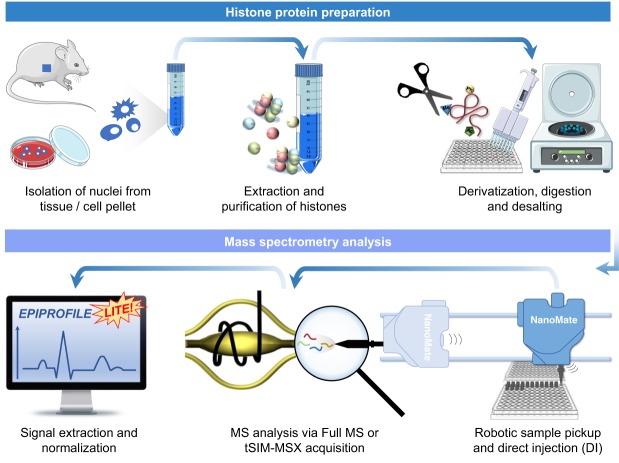
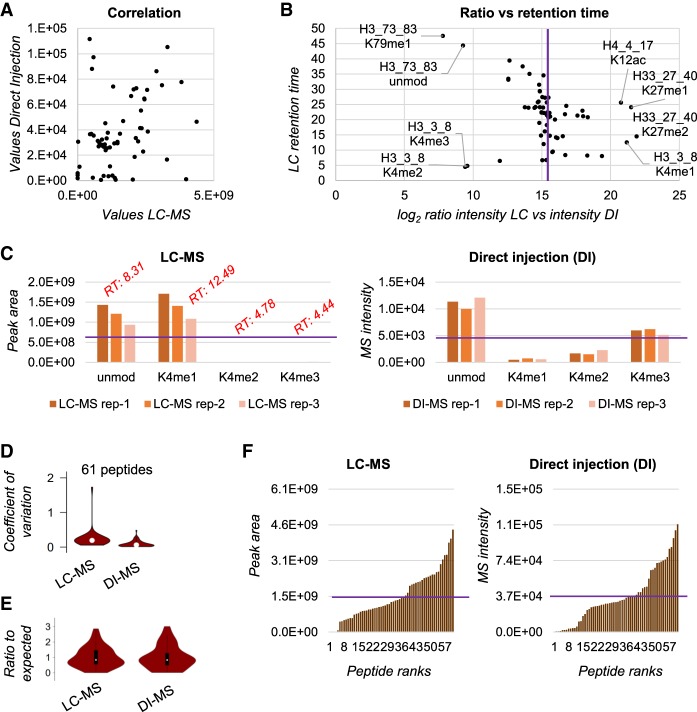
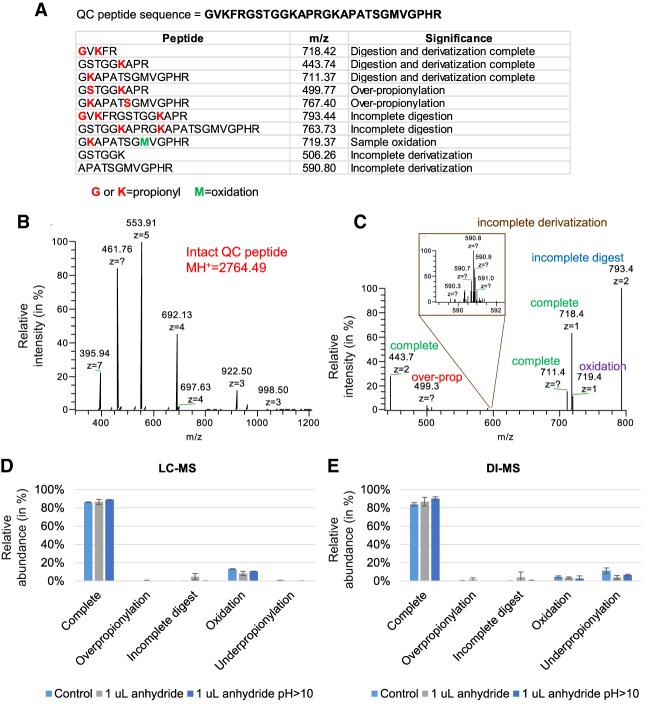
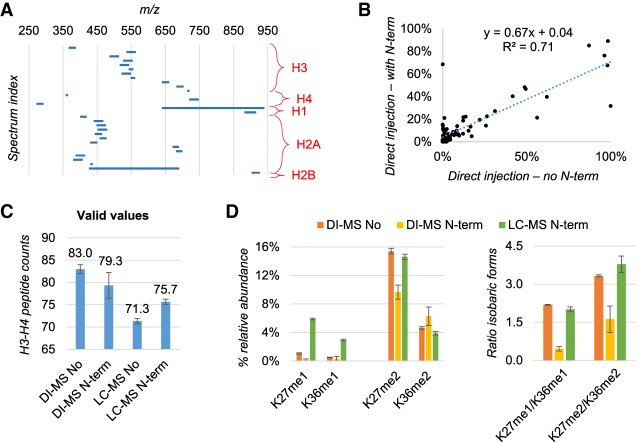
DNA and histone proteins define the structure and composition of chromatin. Histone posttranslational modifications (PTMs) are covalent chemical groups capable of modeling chromatin accessibility, mostly due to their ability in recruiting enzymes responsible for DNA readout and remodeling. Mass spectrometry (MS)-based proteomics is the methodology of choice for large-scale identification and quantification of protein PTMs, including histones. High sensitivity proteomics requires online MS coupling with relatively low throughput and poorly robust nano-liquid chromatography (nanoLC) and, for histone proteins, a 2-d sample preparation that includes histone purification, derivatization, and digestion. We present a new protocol that achieves quantitative data on about 200 histone PTMs from tissue or cell lines in 7 h from start to finish. This protocol includes 4 h of histone extraction, 3 h of derivatization and digestion, and only 1 min of MS analysis via direct injection (DI-MS). We demonstrate that this sample preparation can be parallelized for 384 samples by using multichannel pipettes and 96-well plates. We also engineered the sequence of a synthetic "histone-like" peptide to spike into the sample, of which derivatization and digestion benchmarks the quality of the sample preparation. We ensure that DI-MS does not introduce biases in histone peptide ionization as compared to nanoLC-MS/MS by producing and analyzing a library of synthetically modified histone peptides mixed in equal molarity. Finally, we introduce EpiProfileLite for comprehensive analysis of this new data type. Altogether, our workflow is suitable for high-throughput screening of >1000 samples per day using a single mass spectrometer.
© 2019 Sidoli et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
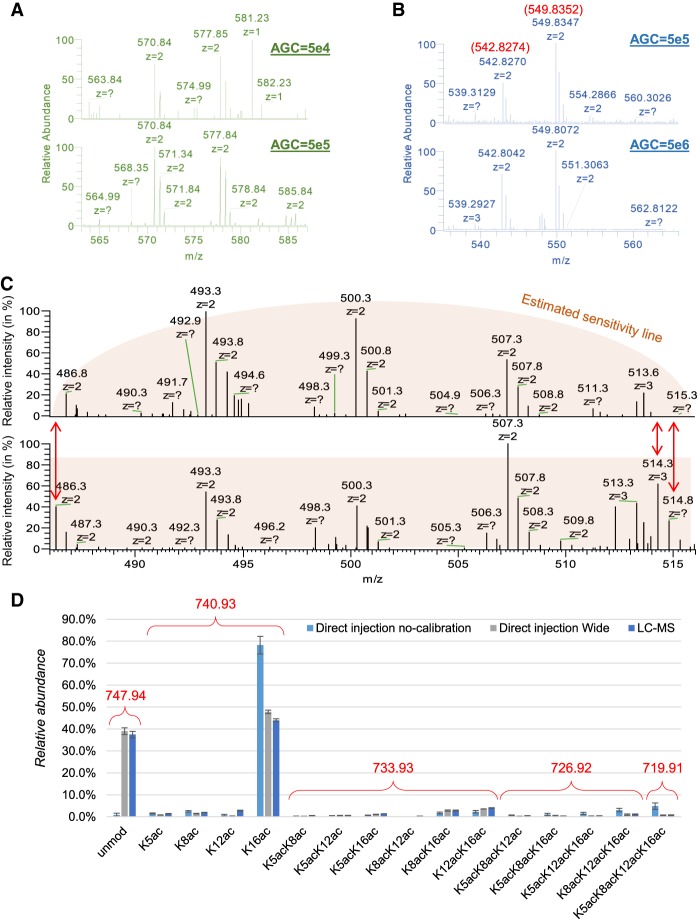
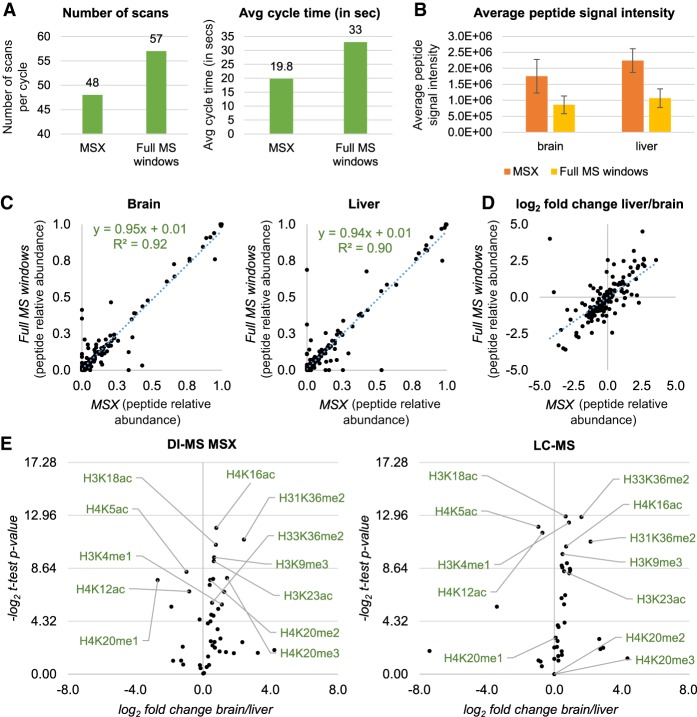
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous