

Bacterial pseudokinase catalyzes protein polyglutamylation to inhibit the SidE-family ubiquitin ligases
- PMID: 31123136
- PMCID: PMC6767918
- DOI: 10.1126/science.aaw7446
Bacterial pseudokinase catalyzes protein polyglutamylation to inhibit the SidE-family ubiquitin ligases
Abstract
Enzymes with a protein kinase fold transfer phosphate from adenosine 5'-triphosphate (ATP) to substrates in a process known as phosphorylation. Here, we show that the Legionella meta-effector SidJ adopts a protein kinase fold, yet unexpectedly catalyzes protein polyglutamylation. SidJ is activated by host-cell calmodulin to polyglutamylate the SidE family of ubiquitin (Ub) ligases. Crystal structures of the SidJ-calmodulin complex reveal a protein kinase fold that catalyzes ATP-dependent isopeptide bond formation between the amino group of free glutamate and the γ-carboxyl group of an active-site glutamate in SidE. We show that SidJ polyglutamylation of SidE, and the consequent inactivation of Ub ligase activity, is required for successful Legionella replication in a viable eukaryotic host cell.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
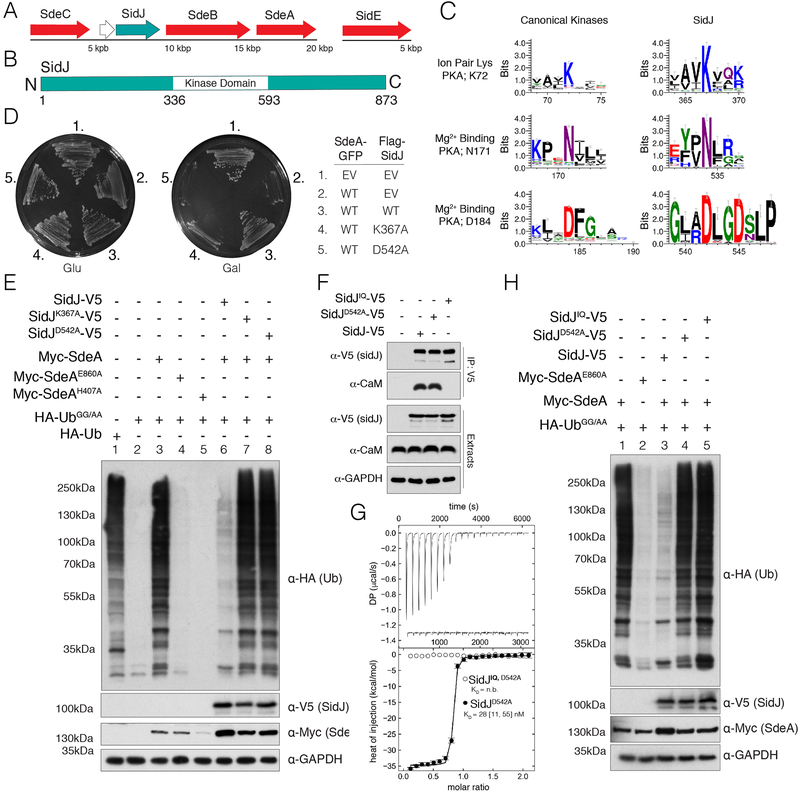
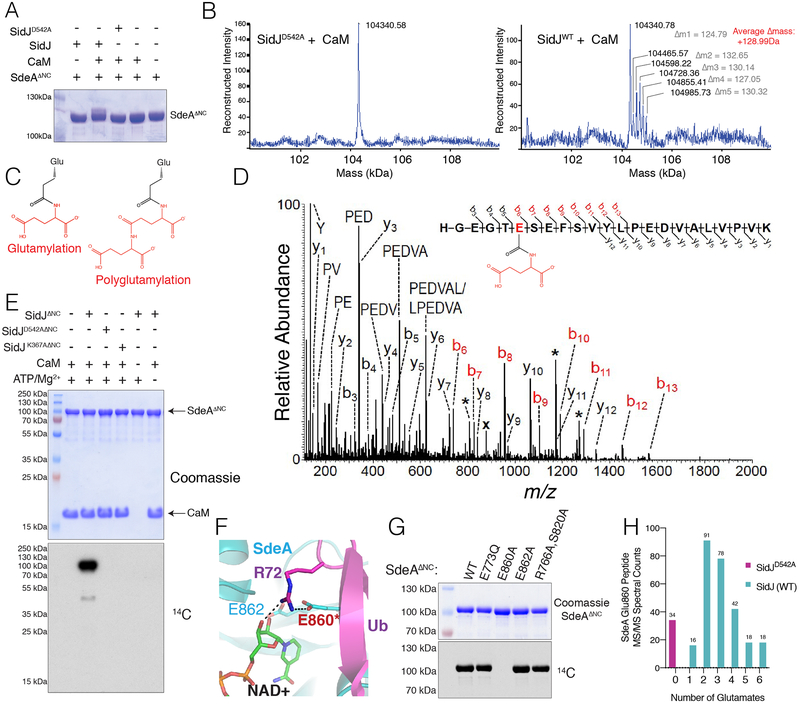
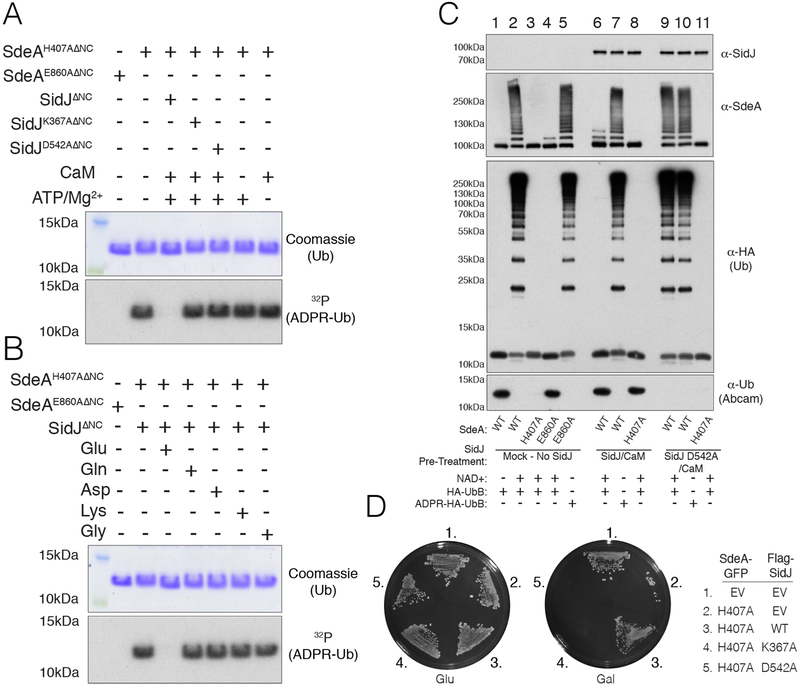
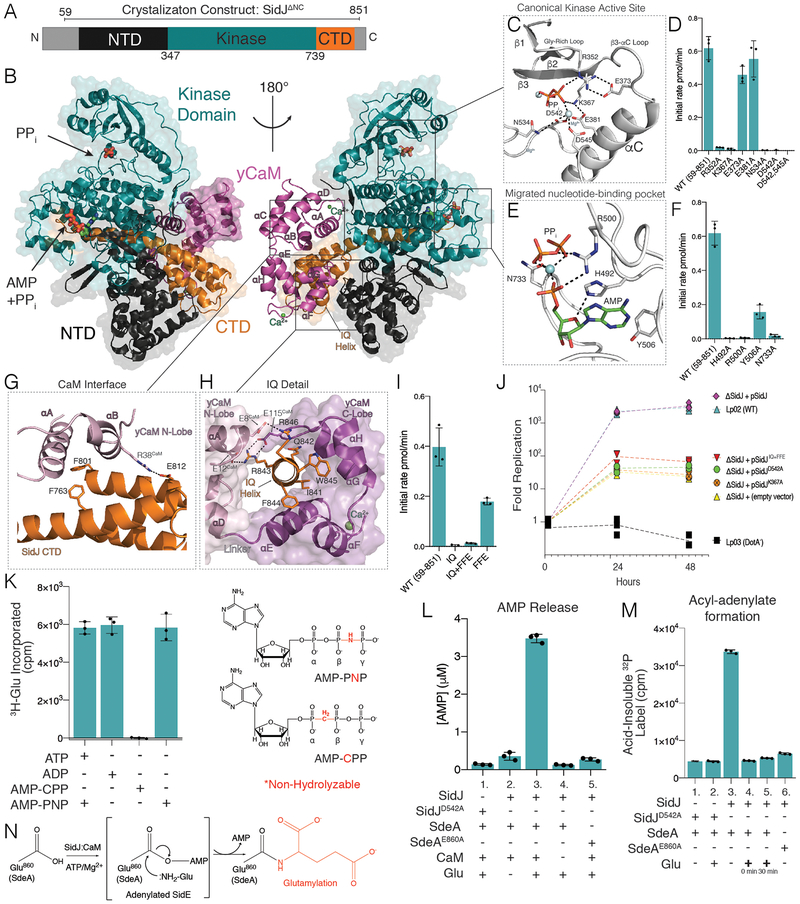
Comment in
-
Glutamylation of Bacterial Ubiquitin Ligases by a Legionella Pseudokinase.Trends Microbiol. 2019 Dec;27(12):967-969. doi: 10.1016/j.tim.2019.09.001. Epub 2019 Oct 14. Trends Microbiol. 2019. PMID: 31624006 Free PMC article.
References
-
- Sherwood RK, Roy CR, Autophagy Evasion and Endoplasmic Reticulum Subversion: The Yin and Yang of Legionella Intracellular Infection. Annu Rev Microbiol 70, 413–433 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
