

Engineering of anti-human interleukin-4 receptor alpha antibodies with potent antagonistic activity
- PMID: 31123339
- PMCID: PMC6533264
- DOI: 10.1038/s41598-019-44253-9
Engineering of anti-human interleukin-4 receptor alpha antibodies with potent antagonistic activity
Abstract
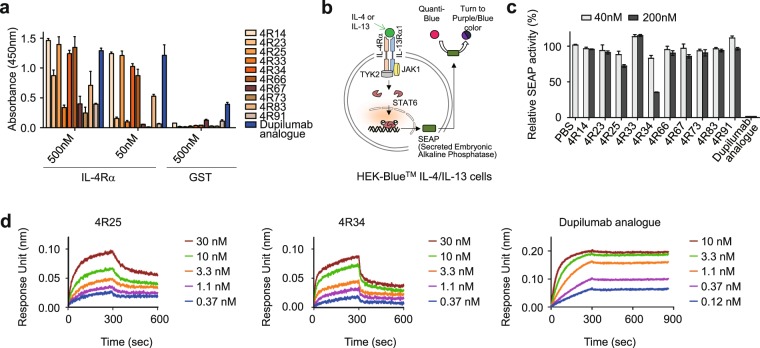
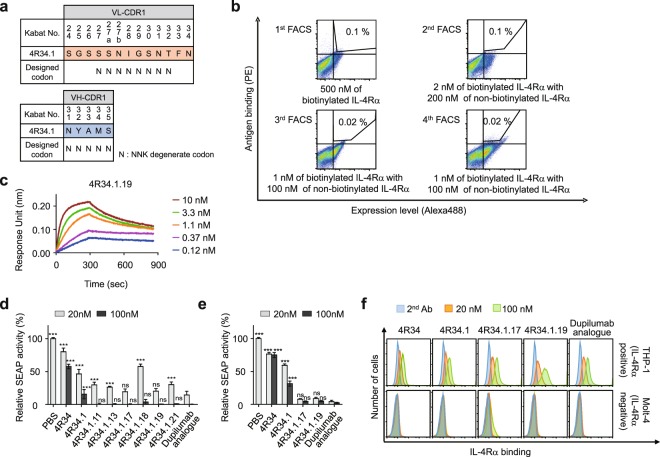
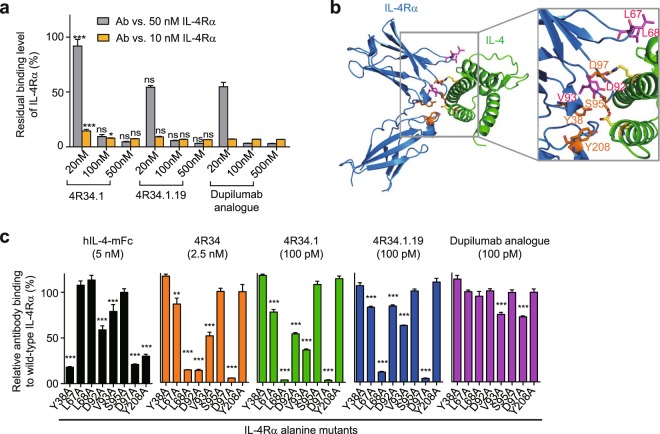
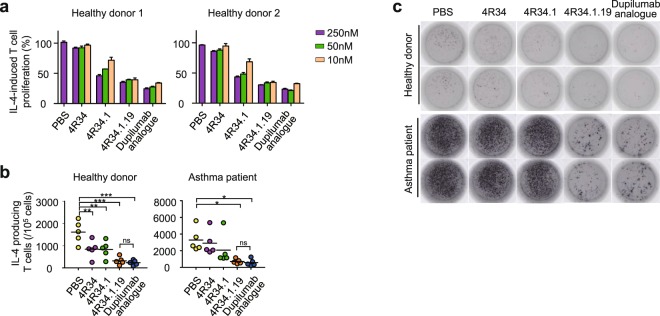
Development of antagonistic antibody (Ab) against interleukin-4 receptor alpha (IL-4Rα) subunit of IL-4/IL-13 receptors is a promising therapeutic strategy for T helper 2 (TH2)-mediated allergic diseases such as asthma and atopic dermatitis. Here we isolated anti-human IL-4Rα antagonistic Abs from a large yeast surface-displayed human Ab library and further engineered their complementarity-determining regions to improve the affinity using yeast display technology, finally generating a candidate Ab, 4R34.1.19. When reformatted as human IgG1 form, 4R34.1.19 specifically bound to IL-4Rα with a high affinity (KD ≈ 178 pM) and effectively blocked IL-4- and IL-13-dependent signaling in a reporter cell system at a comparable level to that of the clinically approved anti-IL-4Rα dupilumab Ab analogue. Epitope mapping by alanine scanning mutagenesis revealed that 4R34.1.19 mainly bound to IL-4 binding sites on IL-4Rα with different epitopes from those of dupilumab analogue. Further, 4R34.1.19 efficiently inhibited IL-4-dependent proliferation of T cells among human peripheral blood mononuclear cells and suppressed the differentiation of naïve CD4+ T cells from healthy donors and asthmatic patients into TH2 cells, the activities of which were comparable to those of dupilumab analogue. Our work demonstrates that both affinity and epitope are critical factors for the efficacy of anti-IL-4Rα antagonistic Abs.
Conflict of interest statement
Y.S.K., H.S.P., J.E.K. and J.K. are listed as inventors on pending patent applications (KR2018-0137199) for antibodies against IL-4Rα described in this work. J.A.K. and S.H.K. declare no financial competing interests. All authors declare no non-financial competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
