

Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6
- PMID: 31123718
- PMCID: PMC6529520
- DOI: 10.1038/s42003-019-0431-5
Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6
Abstract
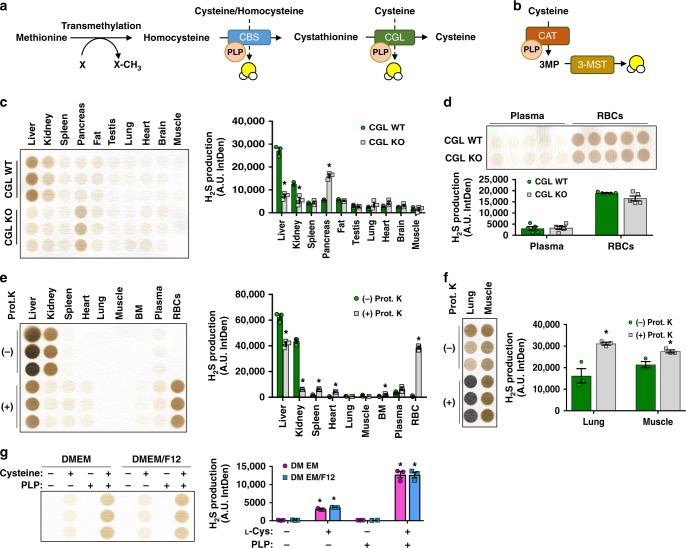
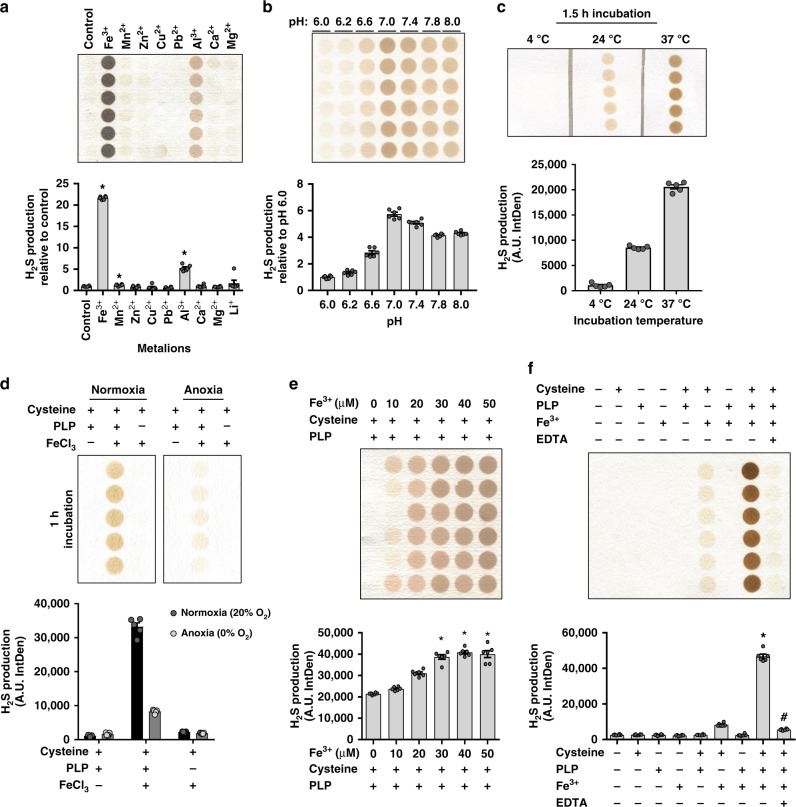
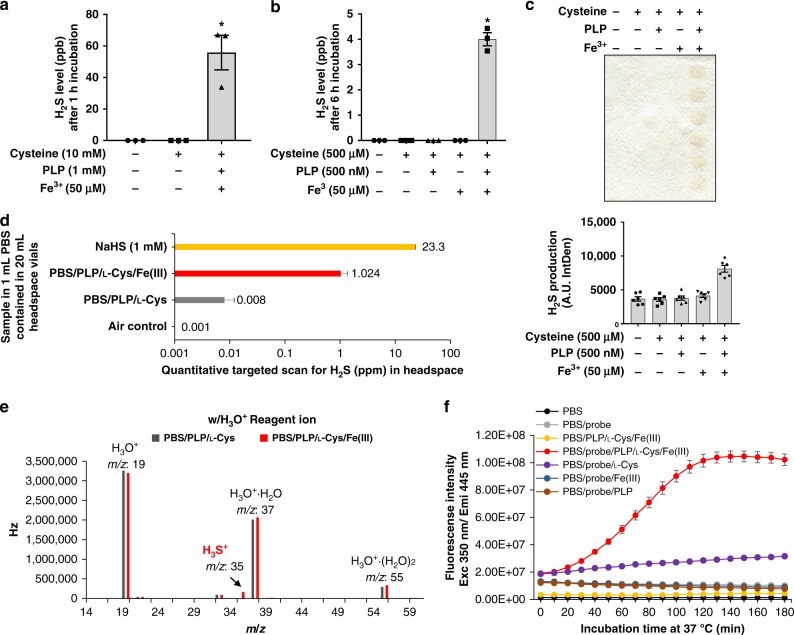
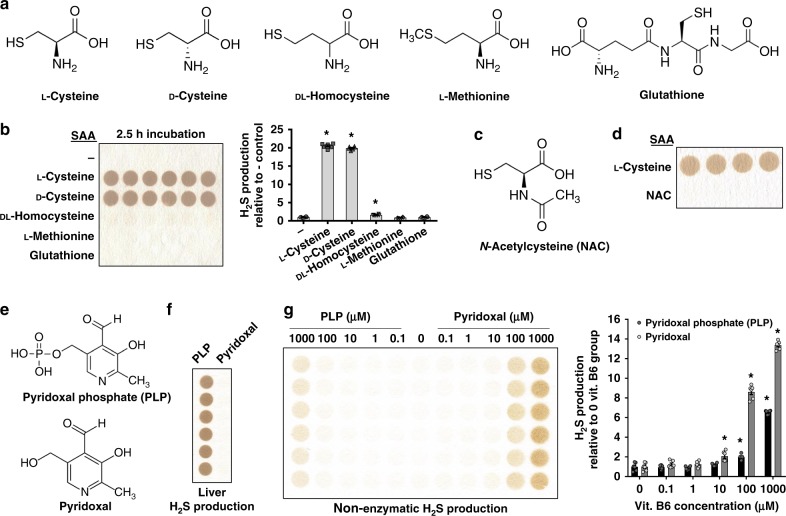
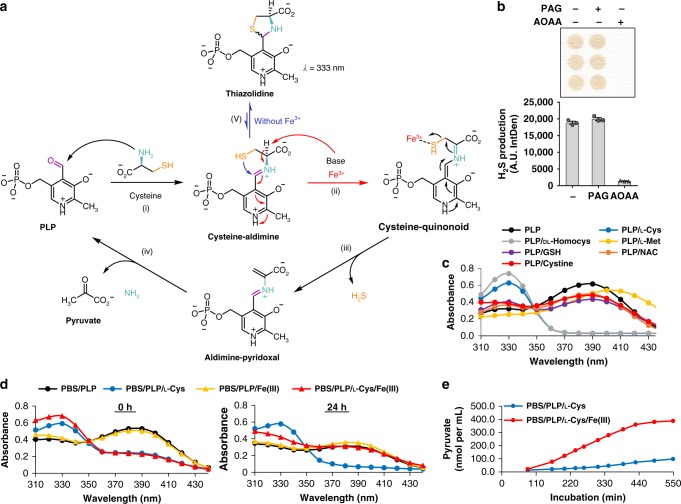
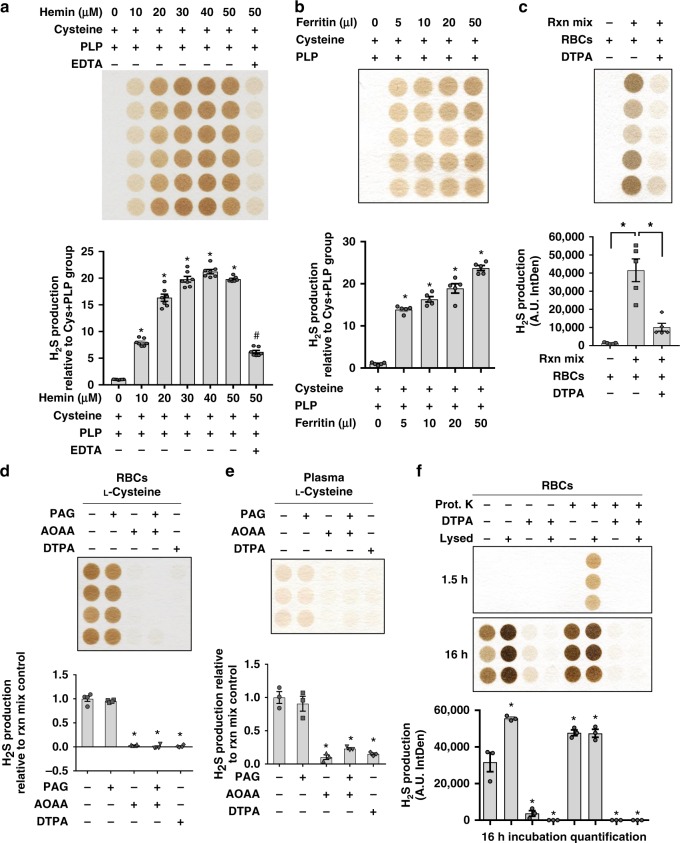
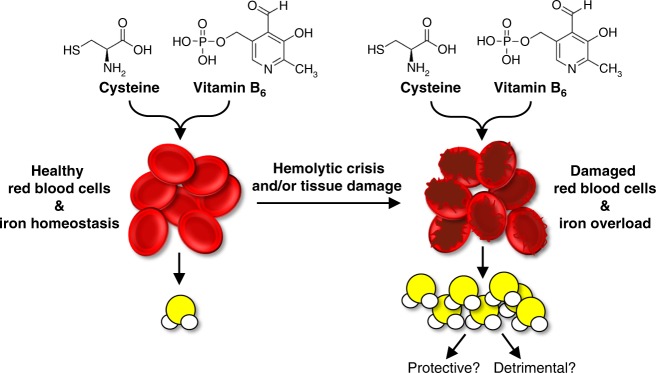
Hydrogen sulfide (H2S) plays important roles in metabolism and health. Its enzymatic generation from sulfur-containing amino acids (SAAs) is well characterized. However, the existence of non-enzymatic H2S production from SAAs, the chemical mechanism, and its biological implications remain unclear. Here we present non-enzymatic H2S production in vitro and in blood via a reaction specific for the SAA cysteine serving as substrate and requires coordinated catalysis by Vitamin B6, pyridoxal(phosphate), and iron under physiological conditions. An initial cysteine-aldimine is formed by nucleophilic attack of the cysteine amino group to the pyridoxal(phosphate) aldehyde group. Free or heme-bound iron drives the formation of a cysteine-quinonoid, thiol group elimination, and hydrolysis of the desulfurated aldimine back to pyridoxal(phosphate). The reaction ultimately produces pyruvate, NH3, and H2S. This work highlights enzymatic production is inducible and robust in select tissues, whereas iron-catalyzed production contributes underappreciated basal H2S systemically with pathophysiological implications in hemolytic, iron overload, and hemorrhagic disorders.
Keywords: Biochemistry; Haematological diseases; Physiology.
Conflict of interest statement
Competing interestsThe authors declare no competing interests.
Figures
References
-
- Olson KR, Straub KD. The role of hydrogen sulfide in evolution and the evolution of hydrogen sulfide in metabolism and signaling. Physiology (Bethesda) 2016;31:60–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
