

Functional MRI and EEG Index Complementary Attentional Modulations
- PMID: 31127004
- PMCID: PMC6668200
- DOI: 10.1523/JNEUROSCI.2519-18.2019
Functional MRI and EEG Index Complementary Attentional Modulations
Abstract
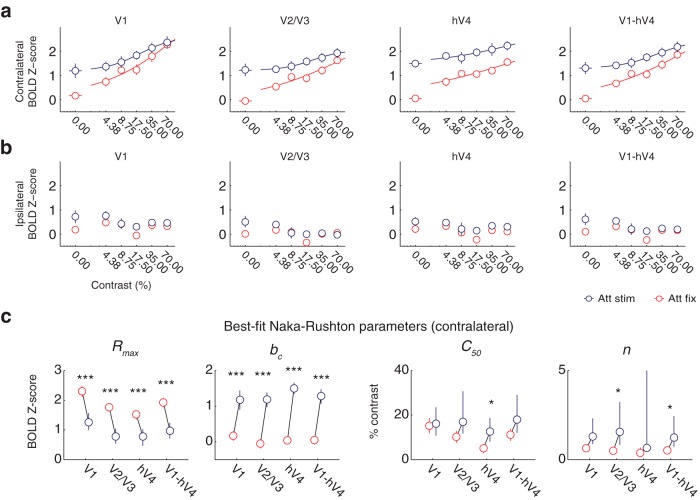
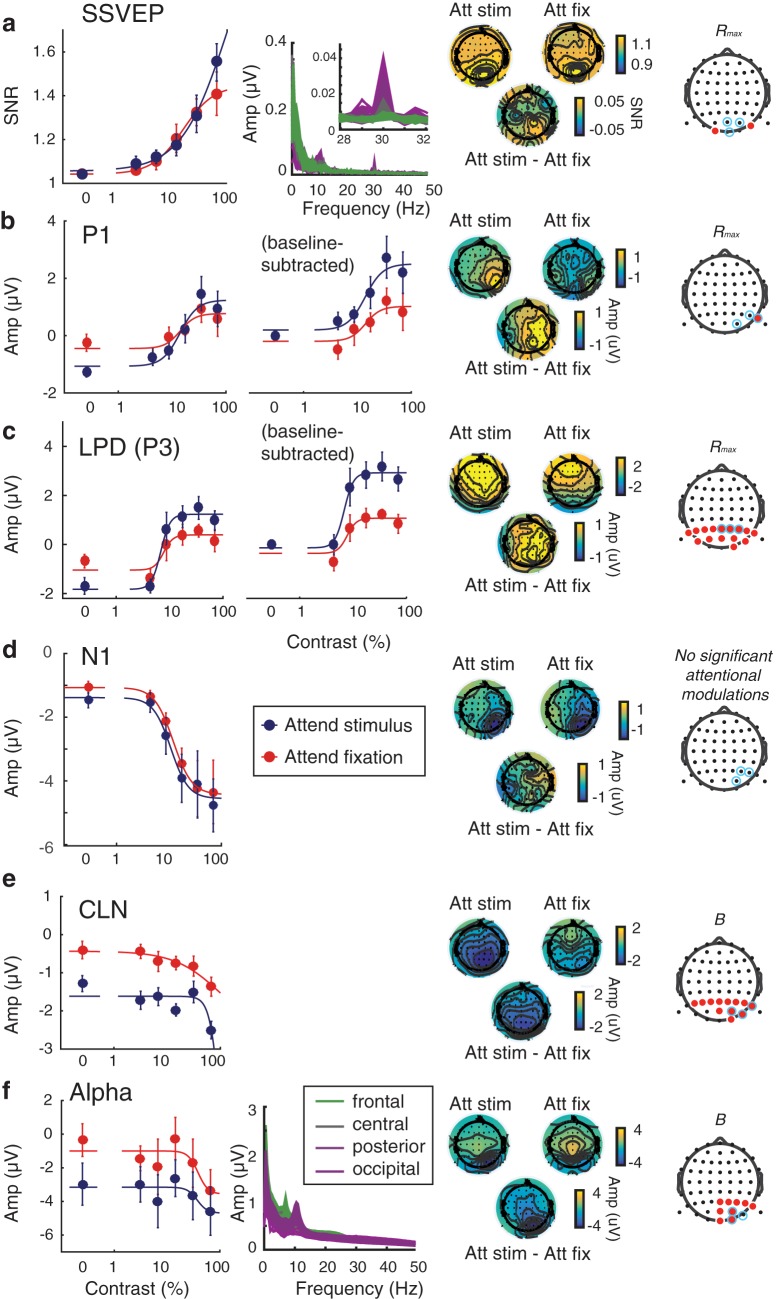
Functional magnetic resonance imaging (fMRI) and electroencephalography (EEG) are two noninvasive methods commonly used to study neural mechanisms supporting visual attention in humans. Studies using these tools, which have complementary spatial and temporal resolutions, implicitly assume they index similar underlying neural modulations related to external stimulus and internal attentional manipulations. Accordingly, they are often used interchangeably for constraining understanding about the impact of bottom-up and top-down factors on neural modulations. To test this core assumption, we simultaneously manipulated bottom-up sensory inputs by varying stimulus contrast and top-down cognitive modulations by changing the focus of spatial attention. Each of the male and female subjects participated in both fMRI and EEG sessions performing the same experimental paradigm. We found categorically different patterns of attentional modulation on fMRI activity in early visual cortex and early stimulus-evoked potentials measured via EEG (e.g., the P1 component and steady-state visually-evoked potentials): fMRI activation scaled additively with attention, whereas evoked EEG components scaled multiplicatively with attention. However, across longer time scales, a contralateral negative-going potential and oscillatory EEG signals in the alpha band revealed additive attentional modulation patterns like those observed with fMRI. These results challenge prior assumptions that fMRI and early stimulus-evoked potentials measured with EEG can be interchangeably used to index the same neural mechanisms of attentional modulations at different spatiotemporal scales. Instead, fMRI measures of attentional modulations are more closely linked with later EEG components and alpha-band oscillations. Considered together, hemodynamic and electrophysiological signals can jointly constrain understanding of the neural mechanisms supporting cognition.SIGNIFICANCE STATEMENT fMRI and EEG have been used as tools to measure the location and timing of attentional modulations in visual cortex and are often used interchangeably for constraining computational models under the assumption that they index similar underlying neural processes. However, by varying attentional and stimulus parameters, we found differential patterns of attentional modulations of fMRI activity in early visual cortex and commonly used stimulus-evoked potentials measured via EEG. Instead, across longer time scales, a contralateral negative-going potential and EEG oscillations in the alpha band exhibited attentional modulations similar to those observed with fMRI. Together, these results suggest that different physiological processes assayed by these complementary techniques must be jointly considered when making inferences about the neural underpinnings of cognitive operations.
Keywords: EEG; attention; contrast response functions; fMRI.
Copyright © 2019 the authors.
Figures





References
-
- Awh E, Jonides J (2001) Overlapping mechanisms of attention and spatial working memory. Trends Cogn Sci 5:119–126. - PubMed
-
- Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc B 57:289–300.