

Recreation of in-host acquired single nucleotide polymorphisms by CRISPR-Cas9 reveals an uncharacterised gene playing a role in Aspergillus fumigatus azole resistance via a non-cyp51A mediated resistance mechanism
- PMID: 31128273
- PMCID: PMC6876285
- DOI: 10.1016/j.fgb.2019.05.005
Recreation of in-host acquired single nucleotide polymorphisms by CRISPR-Cas9 reveals an uncharacterised gene playing a role in Aspergillus fumigatus azole resistance via a non-cyp51A mediated resistance mechanism
Abstract
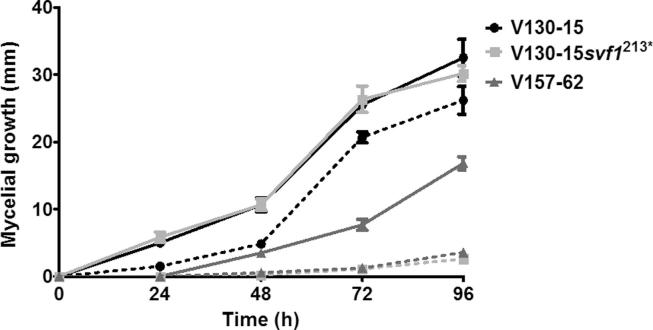
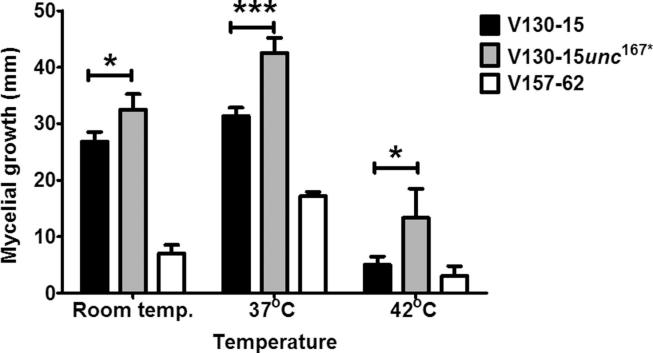
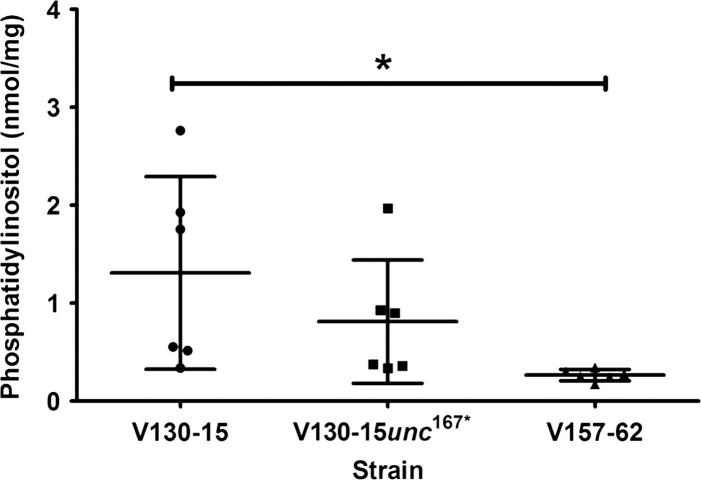
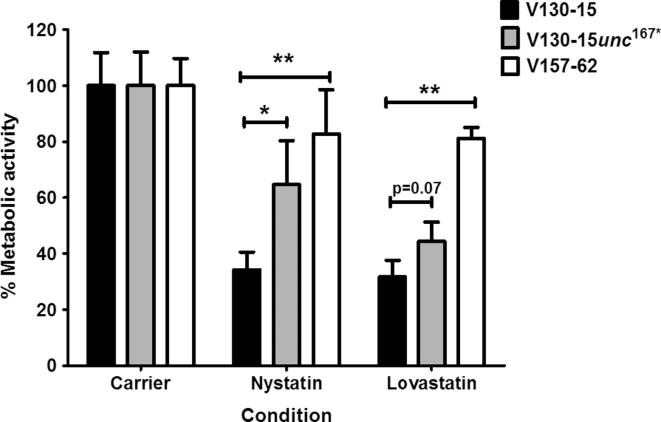
The human host comprises a range of specific niche environments. In order to successfully persist, pathogens such as Aspergillus fumigatus must adapt to these environments. One key example of in-host adaptation is the development of resistance to azole antifungals. Azole resistance in A. fumigatus is increasingly reported worldwide and the most commonly reported mechanisms are cyp51A mediated. Using a unique series of A. fumigatus isolates, obtained from a patient suffering from persistent and recurrent invasive aspergillosis over 2 years, this study aimed to gain insight into the genetic basis of in-host adaptation. Single nucleotide polymorphisms (SNPs) unique to a single isolate in this series, which had developed multi-azole resistance in-host, were identified. Two nonsense SNPs were recreated using CRISPR-Cas9; these were 213* in svf1 and 167* in uncharacterised gene AFUA_7G01960. Phenotypic analyses including antifungal susceptibility testing, mycelial growth rate assessment, lipidomics analysis and statin susceptibility testing were performed to associate genotypes to phenotypes. This revealed a role for svf1 in A. fumigatus oxidative stress sensitivity. In contrast, recapitulation of 167* in AFUA_7G01960 resulted in increased itraconazole resistance. Comprehensive lipidomics analysis revealed decreased ergosterol levels in strains containing this SNP, providing insight to the observed itraconazole resistance. Decreases in ergosterol levels were reflected in increased resistance to lovastatin and nystatin. Importantly, this study has identified a SNP in an uncharacterised gene playing a role in azole resistance via a non-cyp51A mediated resistance mechanism. This mechanism is of clinical importance, as this SNP was identified in a clinical isolate, which acquired azole resistance in-host.
Keywords: Aspergillus fumigatus; Azole resistance; CRISPR-Cas9; Ergosterol; In-host adaptation.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Albarrag A.M., Anderson M.J., Howard S.J., Robson G.D., Warn P.A., Sanglard D., Denning D.W. Interrogation of related clinical pan-azole-resistant Aspergillus fumigatus strains: G138C, Y431C, and G434C single nucleotide polymorphisms in cyp51A, upregulation of cyp51A, and integration and activation of transposon Atf1 in the cyp51A promoter. Antimicrob. Agents Chemother. 2011;55:5113–5121. - PMC - PubMed
-
- Aleksenko A., Clutterbuck A.J. Autonomous Plasmid Replication in Aspergillus nidulans: AMA1 and MATE Elements. Fungal Genet. Biol. 1997;21:373–387. - PubMed
-
- Altschul S.F., Gish W., Miller W., Myers E.W., Lipman D.J. Basic local alignment search tool. J. Mol. Biol. 1990;215:403–410. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
