

Synthetic development: building mammalian multicellular structures with artificial genetic programs
- PMID: 31128430
- PMCID: PMC6778502
- DOI: 10.1016/j.copbio.2019.03.016
Synthetic development: building mammalian multicellular structures with artificial genetic programs
Abstract
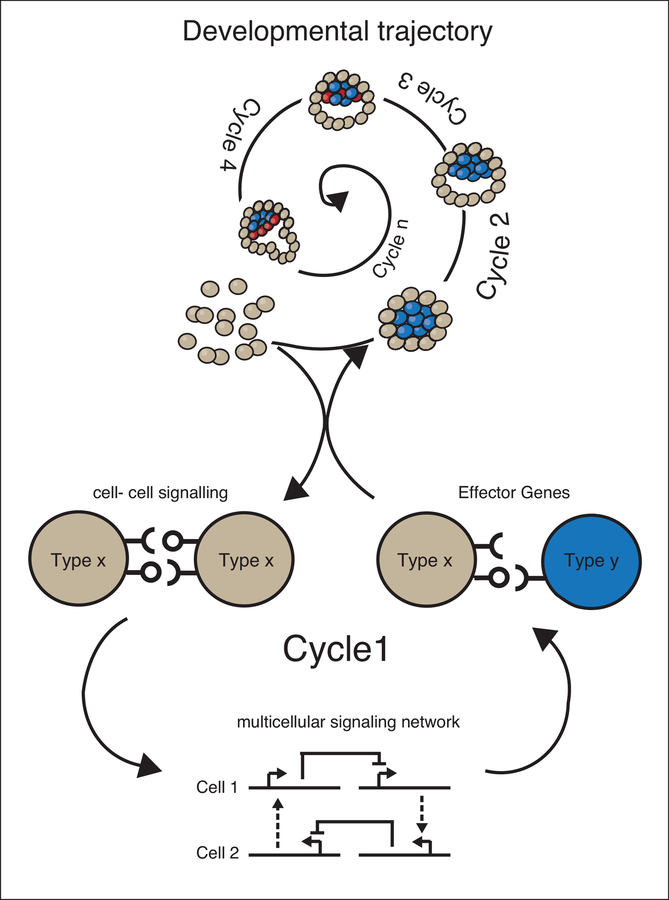
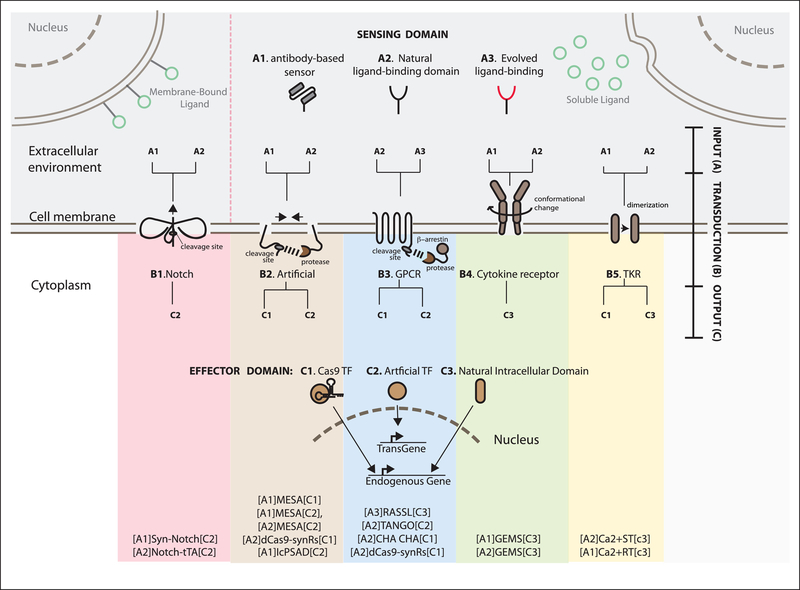
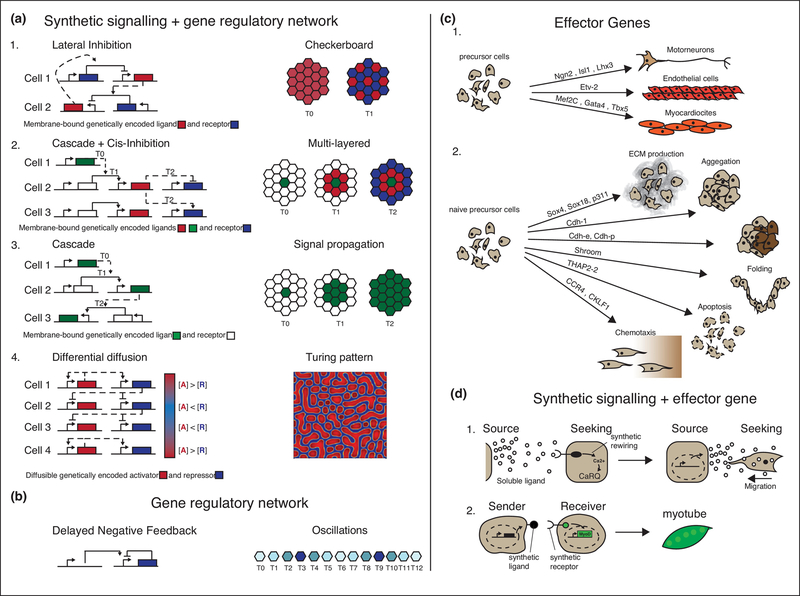
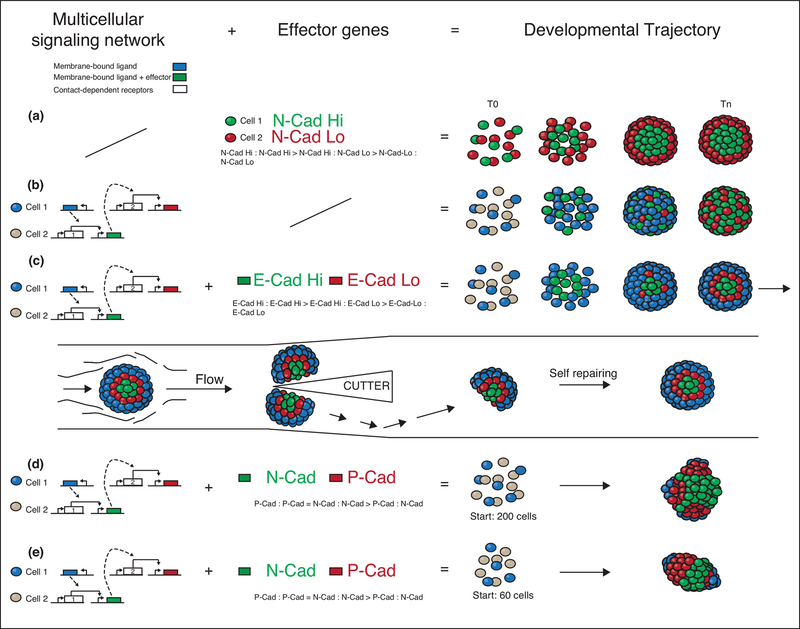
Synthetic biology efforts began in simple single-cell systems, which were relatively easy to manipulate genetically (Cameron et al., 2014). The field grew exponentially in the last two decades, and one of the latest frontiers are synthetic developmental programs for multicellular mammalian systems (Black et al., 2017; Wieland and Fussenegger, 2012) to genetically control features such as patterning or morphogenesis. These programs rely on engineered cell-cell communications, multicellular gene regulatory networks and effector genes. Here, we contextualize the first of these synthetic developmental programs, examine molecular and computational tools that can be used to generate next generation versions, and present the general logic that underpins these approaches. These advances are exciting as they represent a novel way to address both control and understanding in the field of developmental biology and tissue development (Elowitz and Lim, 2010; Velazquez et al., 2018; White et al., 2018; Morsut, 2017). This field is just at the beginning, and it promises to be of major interest in the upcoming years of biomedical research.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
