

Telomere Trimming and DNA Damage as Signatures of High Risk Neuroblastoma
- PMID: 31128432
- PMCID: PMC6535646
- DOI: 10.1016/j.neo.2019.04.002
Telomere Trimming and DNA Damage as Signatures of High Risk Neuroblastoma
Abstract
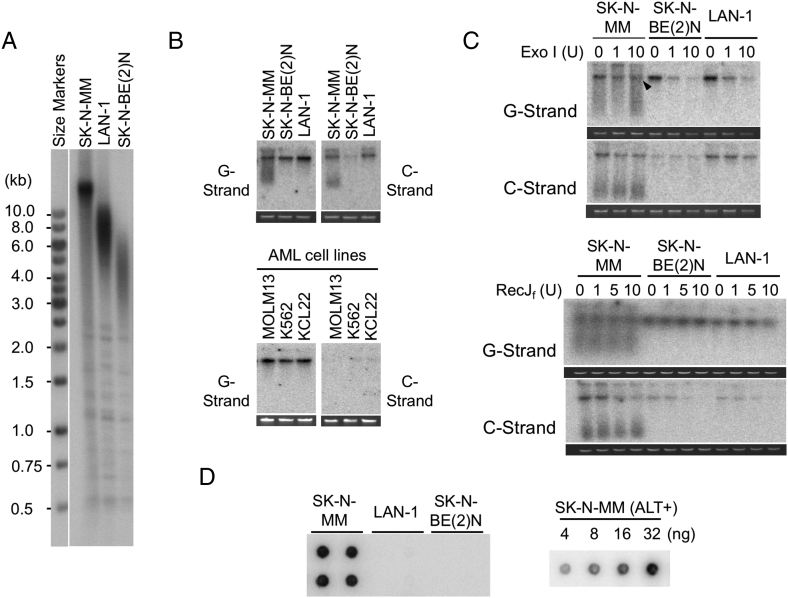
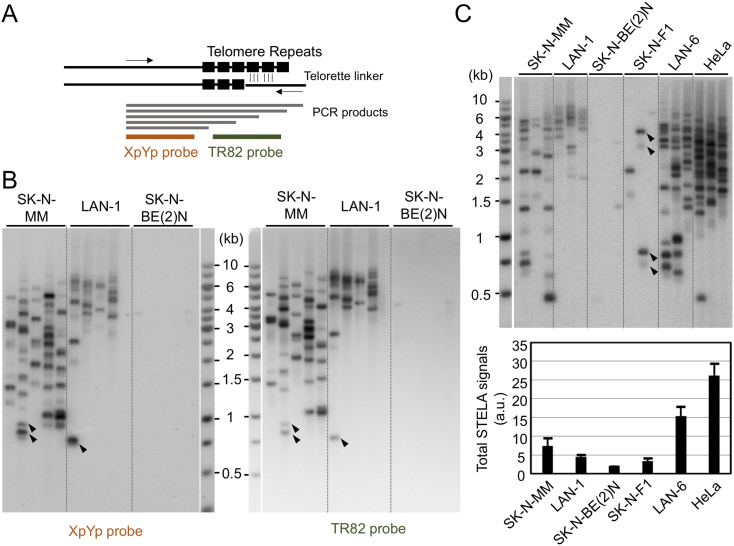
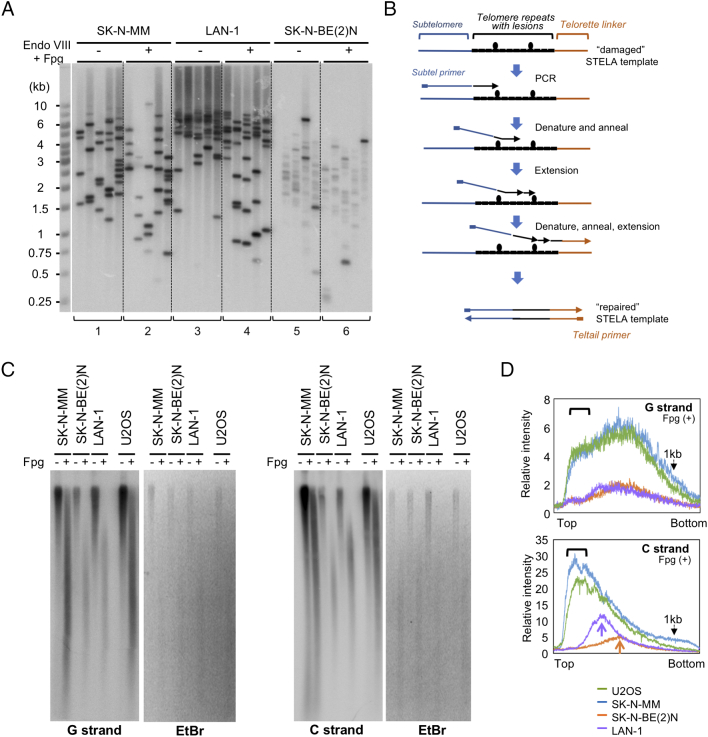
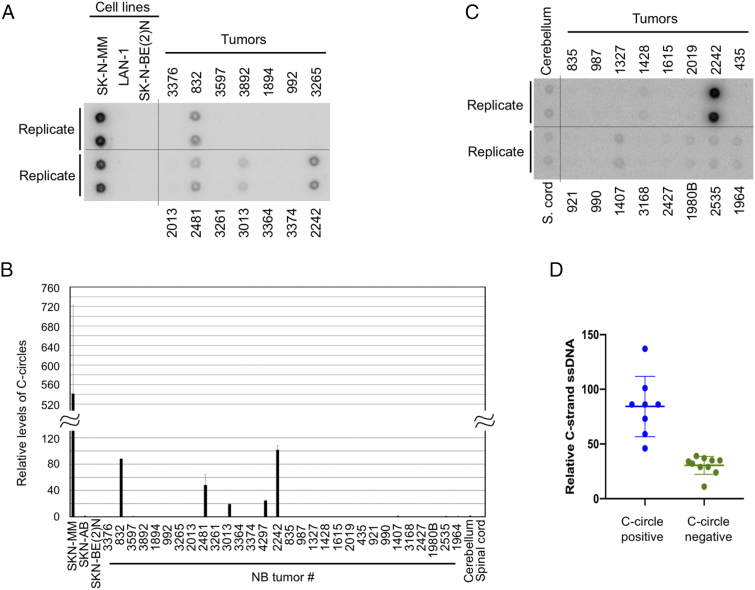
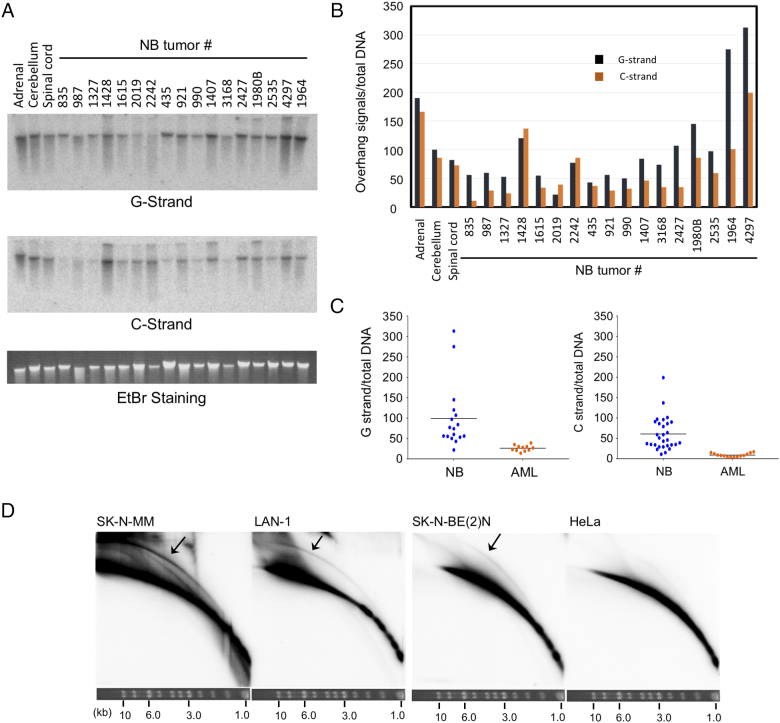
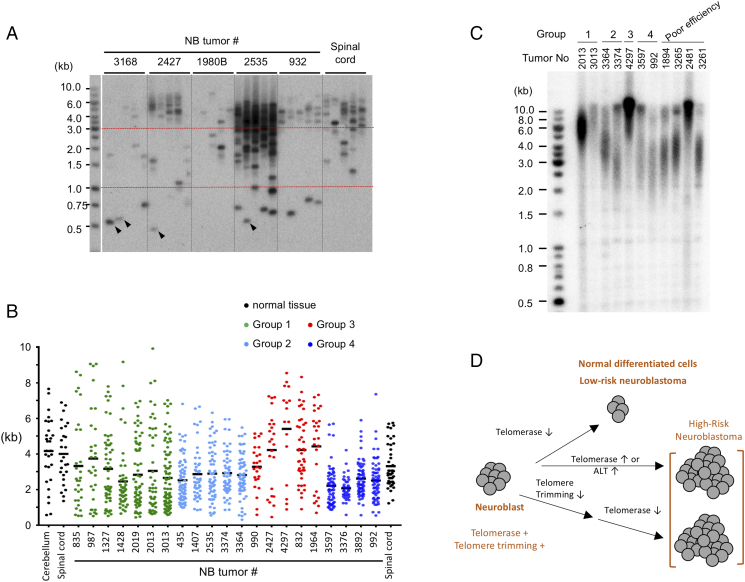
Telomeres play important roles in genome stability and cell proliferation. High risk neuroblastoma (HRNB), an aggressive childhood cancer, is especially reliant on telomere maintenance. Three recurrent genetic aberrations in HRNB (MYCN amplification, TERT re-arrangements, and ATRX mutations) are mutually exclusive and each capable of promoting telomere maintenance mechanisms (i.e., through telomerase or ALT). We analyzed a panel of 5 representative HRNB cell lines and 30 HRNB surgical samples using assays that assess average telomere lengths, length distribution patterns, single-stranded DNA on the G- and C-strand, as well as extra-chromosomal circular telomeres. Our analysis pointed to substantial and variable degrees of telomere DNA damage in HRNB, including pervasive oxidative lesions. Moreover, unlike other cancers, neuroblastoma consistently harbored high levels of C-strand ssDNA overhangs and t-circles, which are consistent with active "telomere trimming". This feature is observed in both telomerase- and ALT-positive tumors and irrespective of telomere length distribution. Moreover, evidence for telomere trimming was detected in normal neural tissues, raising the possibility that TMMs in HRNB evolved in the face of a canonical developmental program of telomere shortening. Telomere trimming by itself appears to distinguish neuroectodermal derived tumors from other human cancers, a distinguishing characteristic with both biologic and therapeutic implications.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
How Do Telomere Abnormalities Regulate the Biology of Neuroblastoma?Biomolecules. 2021 Jul 28;11(8):1112. doi: 10.3390/biom11081112. Biomolecules. 2021. PMID: 34439779 Free PMC article. Review.
-
Telomere Maintenance Mechanisms Define Clinical Outcome in High-Risk Neuroblastoma.Cancer Res. 2020 Jun 15;80(12):2663-2675. doi: 10.1158/0008-5472.CAN-19-3068. Epub 2020 Apr 14. Cancer Res. 2020. PMID: 32291317 Free PMC article.
-
Functional Loss of ATRX and TERC Activates Alternative Lengthening of Telomeres (ALT) in LAPC4 Prostate Cancer Cells.Mol Cancer Res. 2019 Dec;17(12):2480-2491. doi: 10.1158/1541-7786.MCR-19-0654. Epub 2019 Oct 14. Mol Cancer Res. 2019. PMID: 31611308 Free PMC article.
-
Loss of ATRX, genome instability, and an altered DNA damage response are hallmarks of the alternative lengthening of telomeres pathway.PLoS Genet. 2012;8(7):e1002772. doi: 10.1371/journal.pgen.1002772. Epub 2012 Jul 19. PLoS Genet. 2012. PMID: 22829774 Free PMC article.
-
Biological categories of neuroblastoma based on the international neuroblastoma pathology classification for treatment stratification.Pathol Int. 2021 Apr;71(4):232-244. doi: 10.1111/pin.13085. Epub 2021 Mar 3. Pathol Int. 2021. PMID: 33657257 Review.
Cited by
-
Reciprocal impacts of telomerase activity and ADRN/MES differentiation state in neuroblastoma tumor biology.Commun Biol. 2021 Nov 19;4(1):1315. doi: 10.1038/s42003-021-02821-8. Commun Biol. 2021. PMID: 34799676 Free PMC article.
-
Connecting telomere maintenance and regulation to the developmental origin and differentiation states of neuroblastoma tumor cells.J Hematol Oncol. 2022 Aug 27;15(1):117. doi: 10.1186/s13045-022-01337-w. J Hematol Oncol. 2022. PMID: 36030273 Free PMC article. Review.
-
Extrachromosomal Circular DNA: Current Knowledge and Implications for CNS Aging and Neurodegeneration.Int J Mol Sci. 2020 Apr 2;21(7):2477. doi: 10.3390/ijms21072477. Int J Mol Sci. 2020. PMID: 32252492 Free PMC article. Review.
-
Extrachromosomal circular DNA: biogenesis, structure, functions and diseases.Signal Transduct Target Ther. 2022 Oct 2;7(1):342. doi: 10.1038/s41392-022-01176-8. Signal Transduct Target Ther. 2022. PMID: 36184613 Free PMC article. Review.
-
Discovery of a selective TRF2 inhibitor FKB04 induced telomere shortening and senescence in liver cancer cells.Acta Pharmacol Sin. 2024 Jun;45(6):1276-1286. doi: 10.1038/s41401-024-01243-6. Epub 2024 Mar 4. Acta Pharmacol Sin. 2024. PMID: 38438580 Free PMC article.
References
-
- Ward E, DeSantis C, Robbins A, Kohler B, Jemal A. Childhood and adolescent cancer statistics, 2014. CA Cancer J Clin. 2014;64:83–103. - PubMed
-
- Canete A, Gerrard M, Rubie H, Castel V, Di Cataldo A, Munzer C, Ladenstein R, Brichard B, Bermudez JD, Couturier J. Poor survival for infants with MYCN-amplified metastatic neuroblastoma despite intensified treatment: the International Society of Paediatric Oncology European Neuroblastoma Experience. J Clin Oncol. 2009;27:1014–1019. - PubMed
-
- de Lange T. Shelterin-Mediated Telomere Protection. Annu Rev Genet. 2018;52:223–247. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
