

γδTCR-independent origin of neonatal γδ T cells prewired for IL-17 production
- PMID: 31128446
- PMCID: PMC7147991
- DOI: 10.1016/j.coi.2019.04.011
γδTCR-independent origin of neonatal γδ T cells prewired for IL-17 production
Abstract
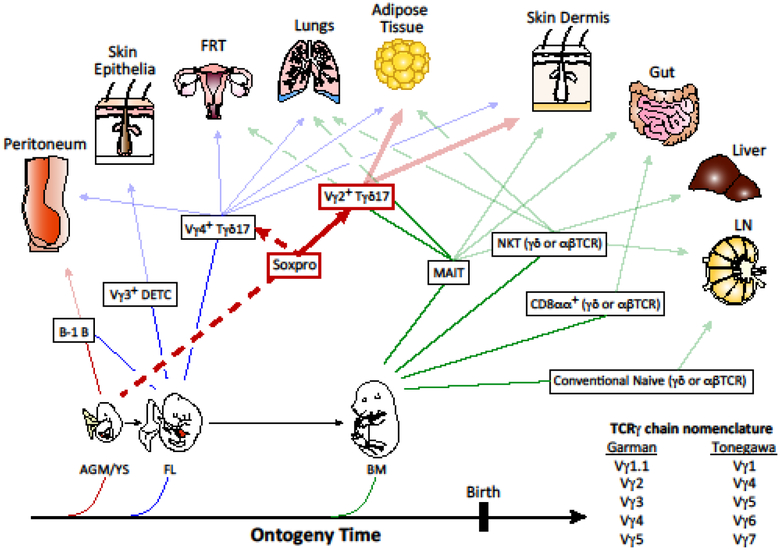
A classical view of T cell lineages consists of two major clades of T cells expressing either the αβ or γδ T cell receptor (TCR). However, genome-wide assessments indicate molecular clusters segregating T cell subsets that are preprogrammed for effector function (innate) from those that mediate conventional adaptive response, regardless of the TCR types. Within this paradigm, γδ T cells remain the prototypic innate-like lymphocytes, many subsets of which are programmed during intrathymic development for committed peripheral tissue localization and effector responses. Emerging evidence for innate γδ T cell lineage choice dictated by developmental gene programs rather than the sensory TCR is discussed in this review.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures

References
-
- Munoz-Ruiz M, Sumaria N, Pennington DJ, Silva-Santos B: Thymic Determinants of gammadelta T Cell Differentiation. Trends Immunol 2017, 38:336–344. - PubMed
-
- Zarin P, Chen EL, In TS, Anderson MK, Zuniga-Pflucker JC: Gamma delta T-cell differentiation and effector function programming, TCR signal strength, when and how much? Cell Immunol 2015, 296:70–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
