

Nucleocytoplasmic O-glycosylation in protists
- PMID: 31128470
- PMCID: PMC6656588
- DOI: 10.1016/j.sbi.2019.03.031
Nucleocytoplasmic O-glycosylation in protists
Abstract
O-Glycosylation is an increasingly recognized modification of intracellular proteins in all kingdoms of life, and its occurrence in protists has been investigated to understand its evolution and its roles in the virulence of unicellular pathogens. We focus here on two kinds of glycoregulation found in unicellular eukaryotes: one is a simple O-fucose modification of dozens if not hundreds of Ser/Thr-rich proteins, and the other a complex pentasaccharide devoted to a single protein associated with oxygen sensing and the assembly of polyubiquitin chains. These modifications are not required for life but contingently modulate biological processes in the social amoeba Dictyostelium and the human pathogen Toxoplasma gondii, and likely occur in diverse unicellular protists. O-Glycosylation that is co-localized in the cytoplasm allows for glycoregulation over the entire life of the protein, contrary to the secretory pathway where glycosylation usually occurs before its delivery to its site of function. Here, we interpret cellular roles of nucleocytoplasmic glycans in terms of current evidence for their effects on the conformation and dynamics of protist proteins, to serve as a guide for future studies to examine their broader significance.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
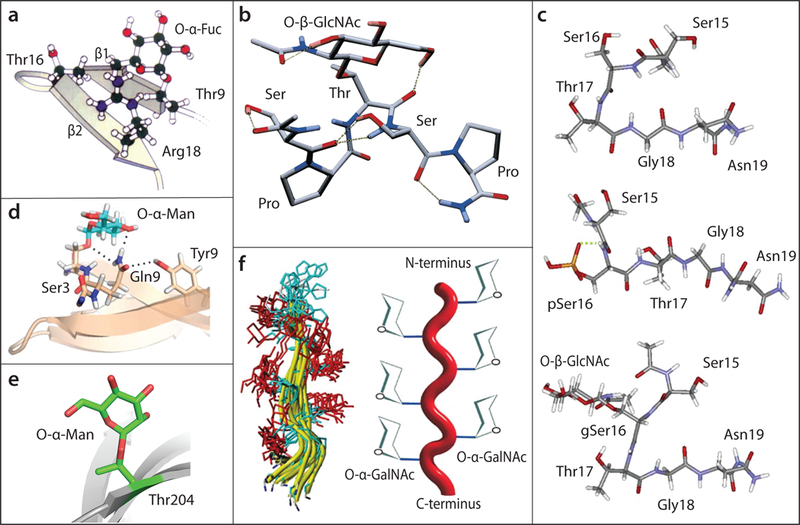
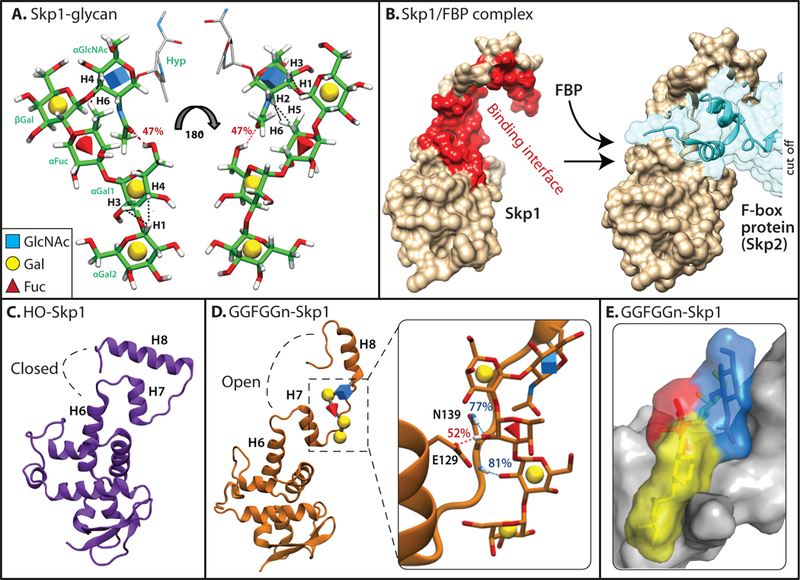
Similar articles
-
Novel antibodies detect nucleocytoplasmic O-fucose in protist pathogens, cellular slime molds, and plants.mSphere. 2025 Feb 25;10(2):e0094524. doi: 10.1128/msphere.00945-24. Epub 2025 Feb 6. mSphere. 2025. PMID: 39912628 Free PMC article.
-
Novel antibodies detect nucleocytoplasmic O-fucose in protist pathogens, cellular slime molds, and plants.bioRxiv [Preprint]. 2024 Oct 22:2024.10.15.618526. doi: 10.1101/2024.10.15.618526. bioRxiv. 2024. Update in: mSphere. 2025 Feb 25;10(2):e0094524. doi: 10.1128/msphere.00945-24. PMID: 39464065 Free PMC article. Updated. Preprint.
-
Evolutionary and functional implications of the complex glycosylation of Skp1, a cytoplasmic/nuclear glycoprotein associated with polyubiquitination.Cell Mol Life Sci. 2003 Feb;60(2):229-40. doi: 10.1007/s000180300018. Cell Mol Life Sci. 2003. PMID: 12678488 Free PMC article. Review.
-
The Skp1 protein from Toxoplasma is modified by a cytoplasmic prolyl 4-hydroxylase associated with oxygen sensing in the social amoeba Dictyostelium.J Biol Chem. 2012 Jul 20;287(30):25098-110. doi: 10.1074/jbc.M112.355446. Epub 2012 May 30. J Biol Chem. 2012. PMID: 22648409 Free PMC article.
-
A cytoplasmic prolyl hydroxylation and glycosylation pathway modifies Skp1 and regulates O2-dependent development in Dictyostelium.Biochim Biophys Acta. 2010 Feb;1800(2):160-71. doi: 10.1016/j.bbagen.2009.11.006. Epub 2009 Nov 13. Biochim Biophys Acta. 2010. PMID: 19914348 Free PMC article. Review.
Cited by
-
Novel antibodies detect nucleocytoplasmic O-fucose in protist pathogens, cellular slime molds, and plants.mSphere. 2025 Feb 25;10(2):e0094524. doi: 10.1128/msphere.00945-24. Epub 2025 Feb 6. mSphere. 2025. PMID: 39912628 Free PMC article.
-
Skp1 Dimerization Conceals Its F-Box Protein Binding Site.Biochemistry. 2020 Apr 21;59(15):1527-1536. doi: 10.1021/acs.biochem.0c00094. Epub 2020 Apr 13. Biochemistry. 2020. PMID: 32227851 Free PMC article.
-
SPINDLY interacts with EIN2 to facilitate ethylene signalling-mediated fruit ripening in tomato.Plant Biotechnol J. 2023 Jan;21(1):219-231. doi: 10.1111/pbi.13939. Epub 2022 Nov 2. Plant Biotechnol J. 2023. PMID: 36204970 Free PMC article.
-
Skp1 isoforms are differentially modified by a dual function prolyl 4-hydroxylase/N-acety lglucosaminyltransferase in a plant pathogen.Glycobiology. 2019 Sep 20;29(10):705-714. doi: 10.1093/glycob/cwz049. Glycobiology. 2019. PMID: 31281925 Free PMC article.
-
A terminal α3-galactose modification regulates an E3 ubiquitin ligase subunit in Toxoplasma gondii.J Biol Chem. 2020 Jul 3;295(27):9223-9243. doi: 10.1074/jbc.RA120.013792. Epub 2020 May 15. J Biol Chem. 2020. PMID: 32414843 Free PMC article.
References
-
- West CM, Hart GW: Nucleocytoplasmic glycosylation. In: Varki A, Cummings RD, Esko JD, Stanley P, Hart GW, Aebi M, Darvill AG, Kinoshita T, Packer NH, Prestegard JH, Schnaar RL, Seeberger PH, editors. Essentials of Glycobiology [Internet] 3rd edition. Cold Spring Harbor Laboratory Press; 2017, Chapter 18
-
- Eustice M, Bond MR, Hanover JA: O-GlcNAc cycling and the regulation of nucleocytoplasmic dynamics. Biochem Soc Trans 2017, 45:427–436. - PubMed
-
- Fredriksen L, Moen A, Adzhubei AA, Mathiesen G, Eijsink VG, Egge-Jacobsen W: Lactobacillus plantarum WCFS1 O-linked protein glycosylation: an extended spectrum of target proteins and modification sites detected by mass spectrometry. Glycobiology 2013, 23:1439–1451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
