

Modulating Hinge Flexibility in the APP Transmembrane Domain Alters γ-Secretase Cleavage
- PMID: 31130234
- PMCID: PMC6554489
- DOI: 10.1016/j.bpj.2019.04.030
Modulating Hinge Flexibility in the APP Transmembrane Domain Alters γ-Secretase Cleavage
Abstract
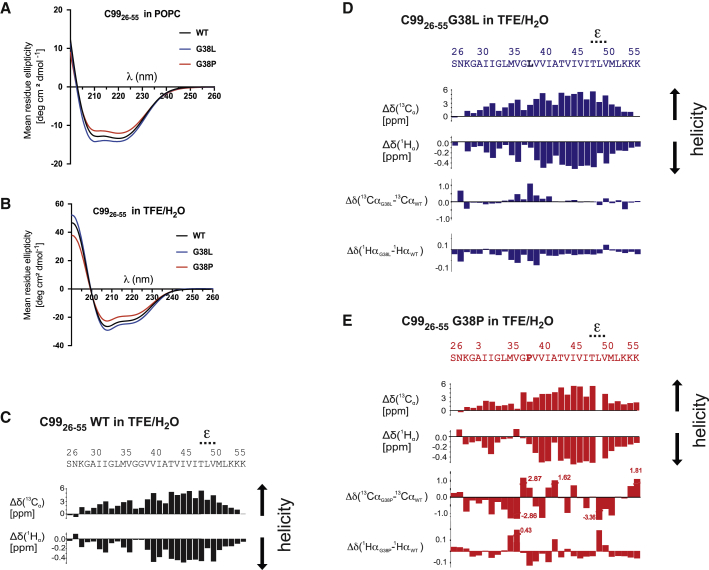
Intramembrane cleavage of the β-amyloid precursor protein C99 substrate by γ-secretase is implicated in Alzheimer's disease pathogenesis. Biophysical data have suggested that the N-terminal part of the C99 transmembrane domain (TMD) is separated from the C-terminal cleavage domain by a di-glycine hinge. Because the flexibility of this hinge might be critical for γ-secretase cleavage, we mutated one of the glycine residues, G38, to a helix-stabilizing leucine and to a helix-distorting proline. Both mutants impaired γ-secretase cleavage and also altered its cleavage specificity. Circular dichroism, NMR, and backbone amide hydrogen/deuterium exchange measurements as well as molecular dynamics simulations showed that the mutations distinctly altered the intrinsic structural and dynamical properties of the substrate TMD. Although helix destabilization and/or unfolding was not observed at the initial ε-cleavage sites of C99, subtle changes in hinge flexibility were identified that substantially affected helix bending and twisting motions in the entire TMD. These resulted in altered orientation of the distal cleavage domain relative to the N-terminal TMD part. Our data suggest that both enhancing and reducing local helix flexibility of the di-glycine hinge may decrease the occurrence of enzyme-substrate complex conformations required for normal catalysis and that hinge mobility can thus be conducive for productive substrate-enzyme interactions.
Copyright © 2019 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures





References
-
- Sun L., Li X., Shi Y. Structural biology of intramembrane proteases: mechanistic insights from rhomboid and S2P to γ-secretase. Curr. Opin. Struct. Biol. 2016;37:97–107. - PubMed
- Sun, L., X. Li, and Y. Shi. 2016. Structural biology of intramembrane proteases: mechanistic insights from rhomboid and S2P to γ-secretase. Curr. Opin. Struct. Biol. 37:97-107. - PubMed
-
- Madala P.K., Tyndall J.D., Fairlie D.P. Update 1 of: proteases universally recognize β strands in their active sites. Chem. Rev. 2010;110:PR1–PR31. - PubMed
- Madala, P. K., J. D. Tyndall, …, D. P. Fairlie. 2010. Update 1 of: proteases universally recognize β strands in their active sites. Chem. Rev. 110:PR1-PR31. - PubMed
-
- Timmer J.C., Zhu W., Salvesen G.S. Structural and kinetic determinants of protease substrates. Nat. Struct. Mol. Biol. 2009;16:1101–1108. - PMC - PubMed
- Timmer, J. C., W. Zhu, …, G. S. Salvesen. 2009. Structural and kinetic determinants of protease substrates. Nat. Struct. Mol. Biol. 16:1101-1108. - PMC - PubMed
-
- Belushkin A.A., Vinogradov D.V., Kazanov M.D. Sequence-derived structural features driving proteolytic processing. Proteomics. 2014;14:42–50. - PMC - PubMed
- Belushkin, A. A., D. V. Vinogradov, …, M. D. Kazanov. 2014. Sequence-derived structural features driving proteolytic processing. Proteomics. 14:42-50. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
