

Polyamines and eIF5A Hypusination Modulate Mitochondrial Respiration and Macrophage Activation
- PMID: 31130465
- PMCID: PMC6688828
- DOI: 10.1016/j.cmet.2019.05.003
Polyamines and eIF5A Hypusination Modulate Mitochondrial Respiration and Macrophage Activation
Abstract
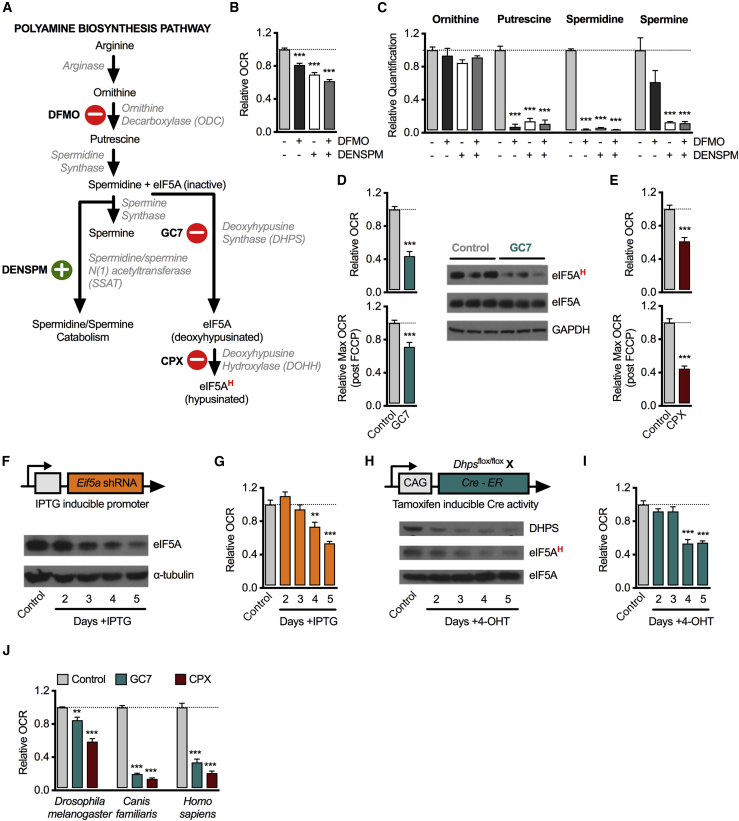
How cells adapt metabolism to meet demands is an active area of interest across biology. Among a broad range of functions, the polyamine spermidine is needed to hypusinate the translation factor eukaryotic initiation factor 5A (eIF5A). We show here that hypusinated eIF5A (eIF5AH) promotes the efficient expression of a subset of mitochondrial proteins involved in the TCA cycle and oxidative phosphorylation (OXPHOS). Several of these proteins have mitochondrial targeting sequences (MTSs) that in part confer an increased dependency on eIF5AH. In macrophages, metabolic switching between OXPHOS and glycolysis supports divergent functional fates stimulated by activation signals. In these cells, hypusination of eIF5A appears to be dynamically regulated after activation. Using in vivo and in vitro models, we show that acute inhibition of this pathway blunts OXPHOS-dependent alternative activation, while leaving aerobic glycolysis-dependent classical activation intact. These results might have implications for therapeutically controlling macrophage activation by targeting the polyamine-eIF5A-hypusine axis.
Keywords: deoxyhypusine hydroxylase; deoxyhypusine synthase; eIF5A; hypusination; immunometabolism; macrophage activation; metabolism; polyamines.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
D.J.P. and E.L.P. have filed a provisional patent application: modulating activation of macrophages (M2 cells) through the polyamine-eIF5A-hypusine axis. E.L.P is a SAB member of ImmunoMet and a founder of Rheos Medicines. E.J.P. is a founder of Rheos Medicines.
Figures




References
-
- Abbruzzese A., Park M.H., Folk J.E. Deoxyhypusine hydroxylase from rat testis. Partial purification and characterization. J. Biol. Chem. 1986;261:3085–3089. - PubMed
-
- Arpa L., Valledor A.F., Lloberas J., Celada A. IL-4 blocks M-CSF-dependent macrophage proliferation by inducing p21WAF1 in a STAT6-dependent way. Eur. J. Immunol. 2009;39:514–526. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
