

Slc12a8 is a nicotinamide mononucleotide transporter
- PMID: 31131364
- PMCID: PMC6530925
- DOI: 10.1038/s42255-018-0009-4
Slc12a8 is a nicotinamide mononucleotide transporter
Erratum in
-
Author Correction: Slc12a8 is a nicotinamide mononucleotide transporter.Nat Metab. 2019 Jul;1(7):743. doi: 10.1038/s42255-019-0088-x. Nat Metab. 2019. PMID: 32694647
Abstract
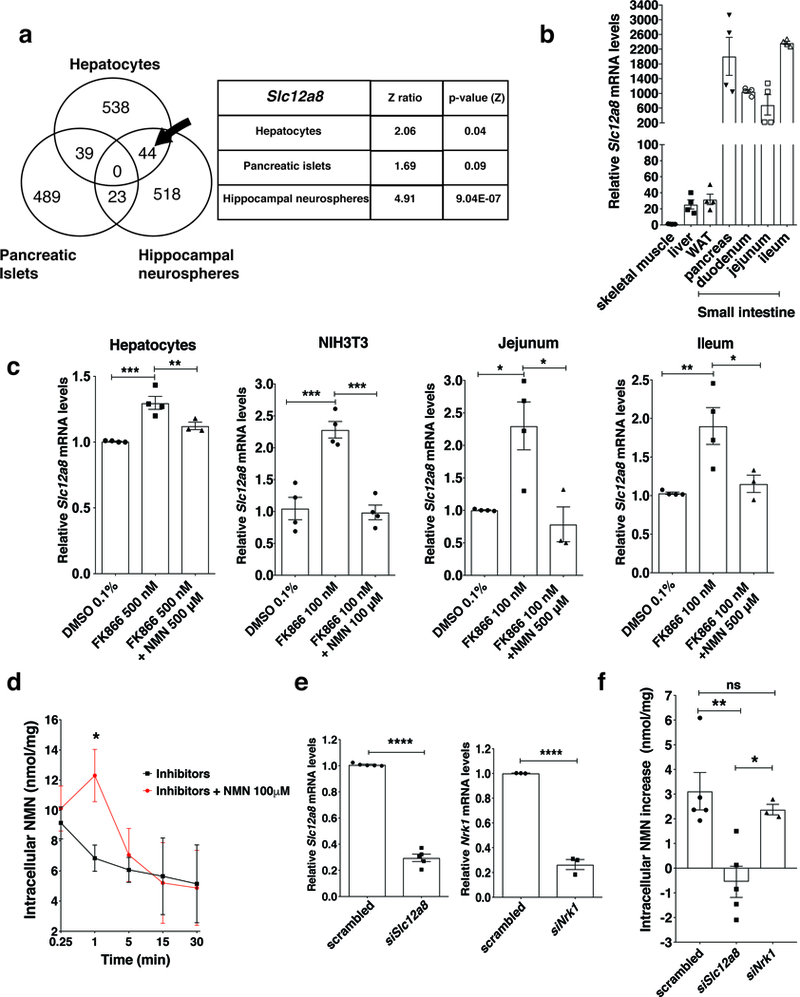
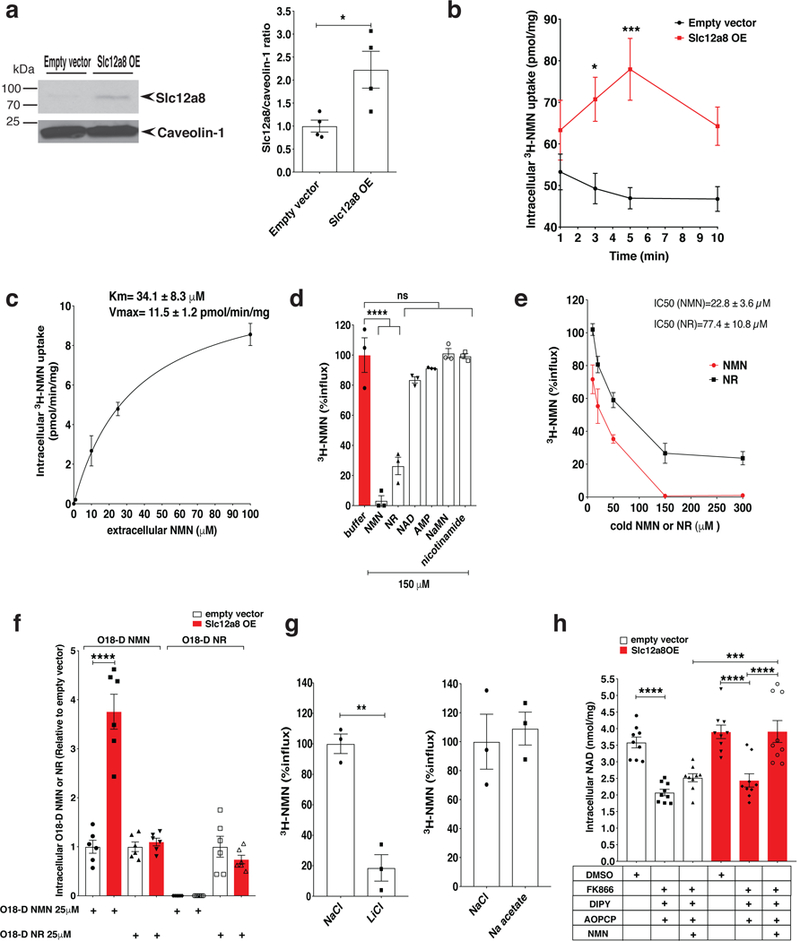
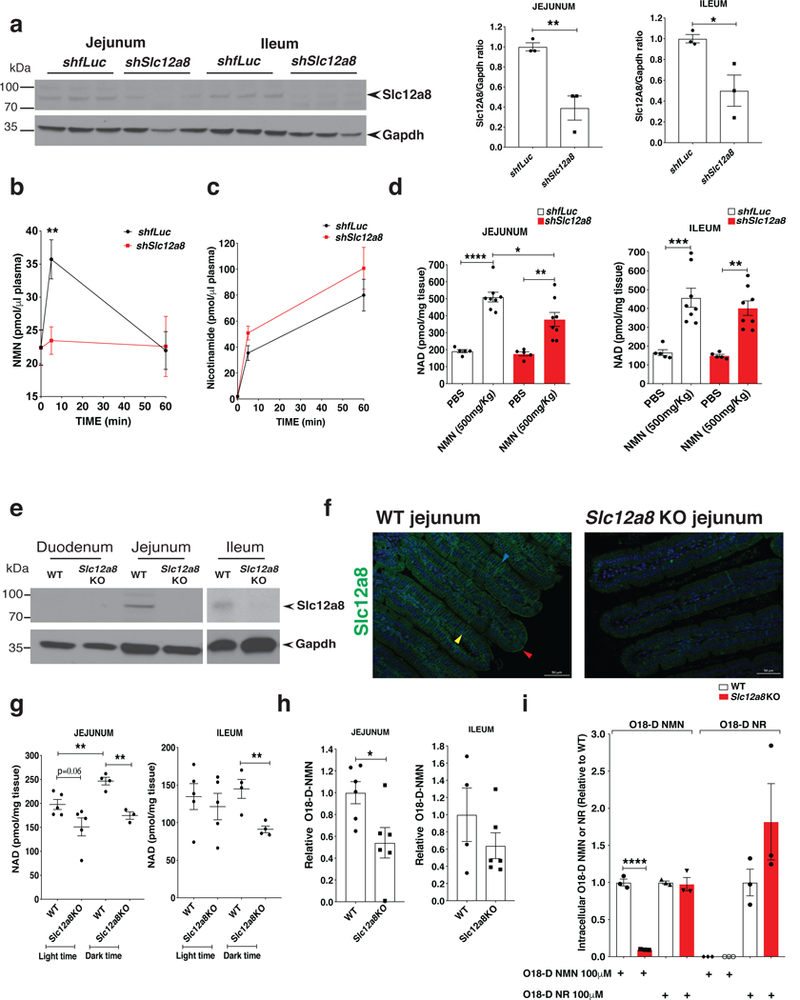
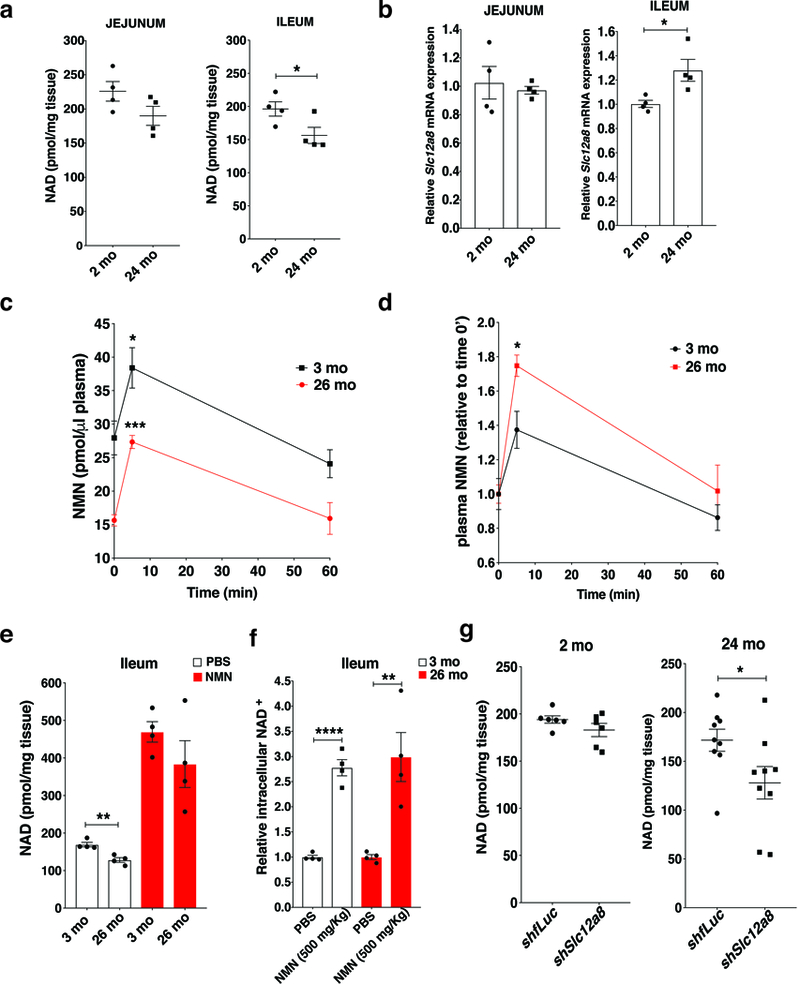
Nicotinamide mononucleotide (NMN) is a biosynthetic precursor of NAD+ known to promote cellular NAD+ production and counteract age-associated pathologies associated with a decline in tissue NAD+ levels. How NMN is taken up into cells has not been entirely clear. Here we show that the Slc12a8 gene encodes a specific NMN transporter. We find that Slc12a8 is highly expressed and regulated by NAD+ in the murine small intestine. Slc12a8 knockdown abrogates the uptake of NMN in vitro and in vivo. We further show that Slc12a8 specifically transports NMN, but not nicotinamide riboside, and that NMN transport depends on the presence of sodium ion. Slc12a8 deficiency significantly decreases NAD+ levels in the jejunum and ileum, which is associated with reduced NMN uptake as traced by doubly labeled isotopic NMN. Finally, we observe that Slc12a8 expression is upregulated in the aged murine ileum, which contributes to the maintenance of ileal NAD+ levels. Our work identifies the first NMN transporter and demonstrates that Slc12a8 has a critical role in regulating intestinal NAD+ metabolism.
Figures
Comment in
-
Absence of evidence that Slc12a8 encodes a nicotinamide mononucleotide transporter.Nat Metab. 2019 Jul;1(7):660-661. doi: 10.1038/s42255-019-0085-0. Epub 2019 Jul 12. Nat Metab. 2019. PMID: 32694648 No abstract available.
-
Reply to: Absence of evidence that Slc12a8 encodes a nicotinamide mononucleotide transporter.Nat Metab. 2019 Jul;1(7):662-665. doi: 10.1038/s42255-019-0086-z. Epub 2019 Jul 12. Nat Metab. 2019. PMID: 32694650 No abstract available.
References
-
- Yoshino J, Mills KF, Yoon MJ & Imai S Nicotinamide mononucleotide, a key NAD(+) intermediate, treats the pathophysiology of diet- and age-induced diabetes in mice. Cell Metab. 14, 528–536, doi:S1550-4131(11)00346-9[pii]10.1016/j.cmet.2011.08.014 (2011). - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
