

Loss of DUX causes minor defects in zygotic genome activation and is compatible with mouse development
- PMID: 31133747
- PMCID: PMC6545155
- DOI: 10.1038/s41588-019-0418-7
Loss of DUX causes minor defects in zygotic genome activation and is compatible with mouse development
Abstract
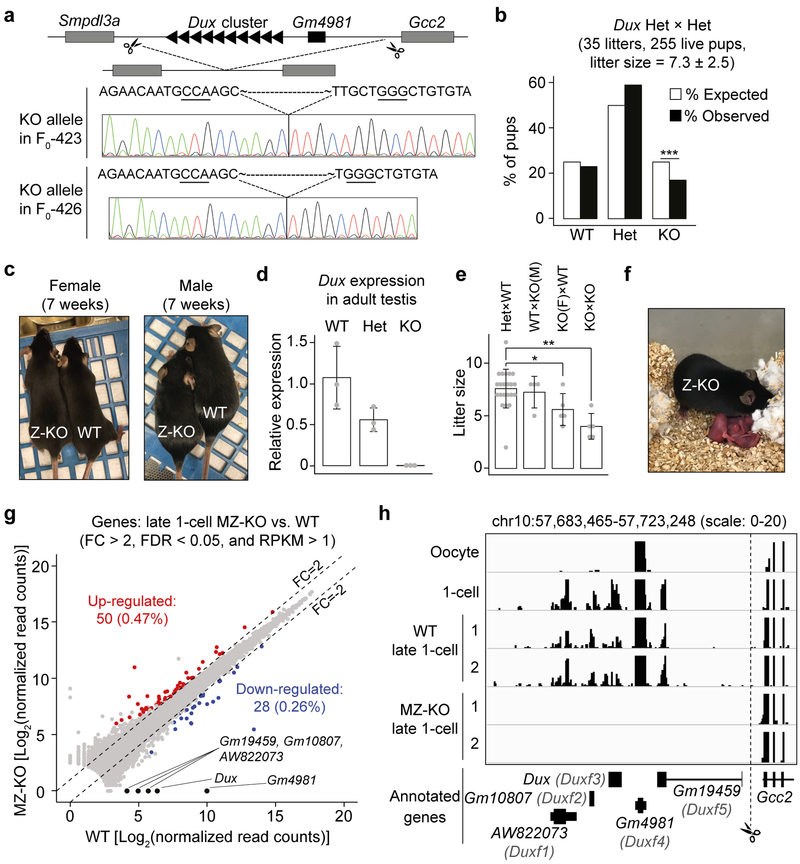
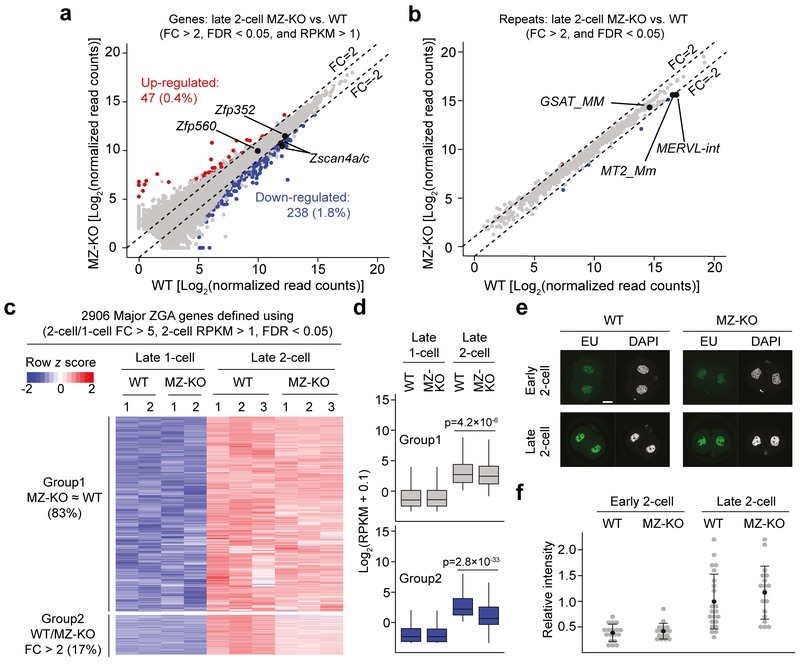
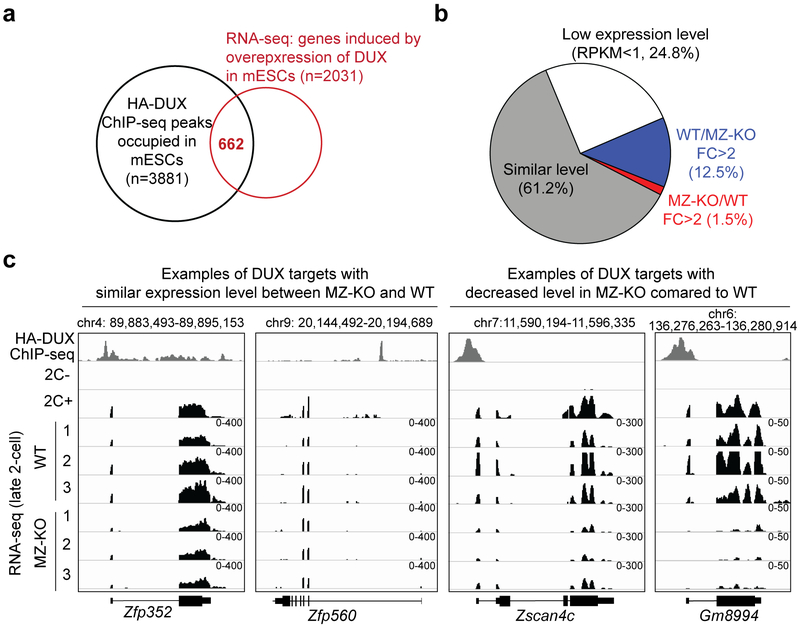
How maternal factors in oocytes trigger zygotic genome activation (ZGA) is a long-standing question in developmental biology. Recent studies in 2-cell-like embryonic stem cells (2C-like cells) suggest that transcription factors of the DUX family are key regulators of ZGA in placental mammals1,2. To characterize the role of DUX in ZGA, we generated Dux cluster knockout (KO) mouse lines. Unexpectedly, we found that both Dux zygotic KO (Z-KO) and maternal and zygotic KO (MZ-KO) embryos can survive to adulthood despite showing reduced developmental potential. Furthermore, transcriptome profiling of the MZ-KO embryos revealed that loss of DUX has minimal effects on ZGA and most DUX targets in 2C-like cells are normally activated in MZ-KO embryos. Thus, contrary to the key function of DUX in inducing 2C-like cells, our data indicate that DUX has only a minor role in ZGA and that loss of DUX is compatible with mouse development.
Figures
References
-
- Hamatani T, Carter MG, Sharov AA & Ko MS Dynamics of global gene expression changes during mouse preimplantation development. Dev Cell 6, 117–31 (2004). - PubMed
-
- Wang QT et al. A genome-wide study of gene activity reveals developmental signaling pathways in the preimplantation mouse embryo. Dev Cell 6, 133–44 (2004). - PubMed
-
- Zeng F, Baldwin DA & Schultz RM Transcript profiling during preimplantation mouse development. Dev Biol 272, 483–96 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
