

Mitochondrial metabolism promotes adaptation to proteotoxic stress
- PMID: 31133756
- PMCID: PMC8183600
- DOI: 10.1038/s41589-019-0291-9
Mitochondrial metabolism promotes adaptation to proteotoxic stress
Erratum in
-
Author Correction: Mitochondrial metabolism promotes adaptation to proteotoxic stress.Nat Chem Biol. 2019 Jul;15(7):757. doi: 10.1038/s41589-019-0315-5. Nat Chem Biol. 2019. PMID: 31164776 Free PMC article.
Abstract
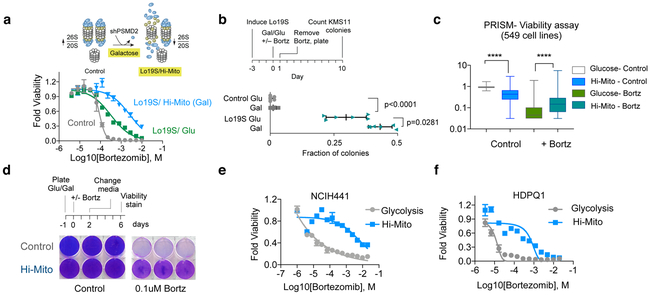
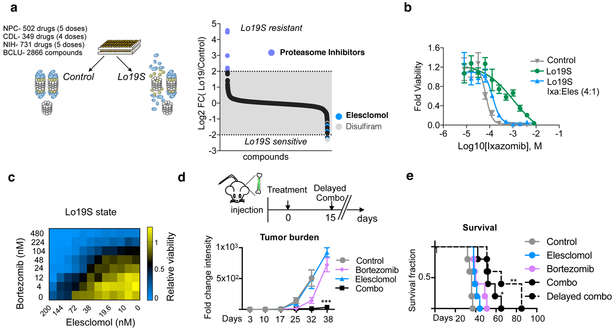
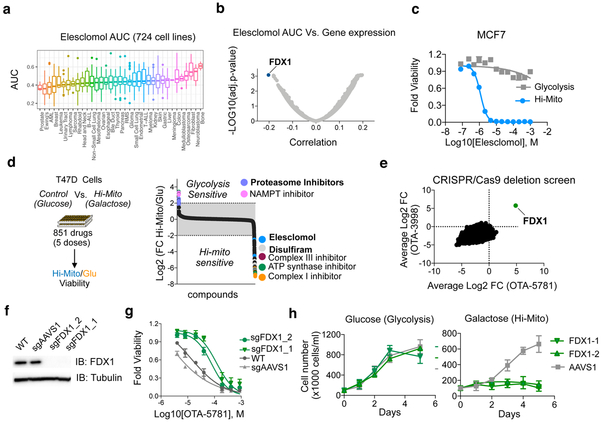
The mechanisms by which cells adapt to proteotoxic stress are largely unknown, but are key to understanding how tumor cells, particularly in vivo, are largely resistant to proteasome inhibitors. Analysis of cancer cell lines, mouse xenografts and patient-derived tumor samples all showed an association between mitochondrial metabolism and proteasome inhibitor sensitivity. When cells were forced to use oxidative phosphorylation rather than glycolysis, they became proteasome-inhibitor resistant. This mitochondrial state, however, creates a unique vulnerability: sensitivity to the small molecule compound elesclomol. Genome-wide CRISPR-Cas9 screening showed that a single gene, encoding the mitochondrial reductase FDX1, could rescue elesclomol-induced cell death. Enzymatic function and nuclear-magnetic-resonance-based analyses further showed that FDX1 is the direct target of elesclomol, which promotes a unique form of copper-dependent cell death. These studies explain a fundamental mechanism by which cells adapt to proteotoxic stress and suggest strategies to mitigate proteasome inhibitor resistance.
Figures



References
-
- Adams J et al. Proteasome inhibitors: a novel class of potent and effective antitumor agents. Cancer Res 59, 2615–2622 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
