

Division plane placement in pleomorphic archaea is dynamically coupled to cell shape
- PMID: 31136034
- PMCID: PMC6736733
- DOI: 10.1111/mmi.14316
Division plane placement in pleomorphic archaea is dynamically coupled to cell shape
Abstract
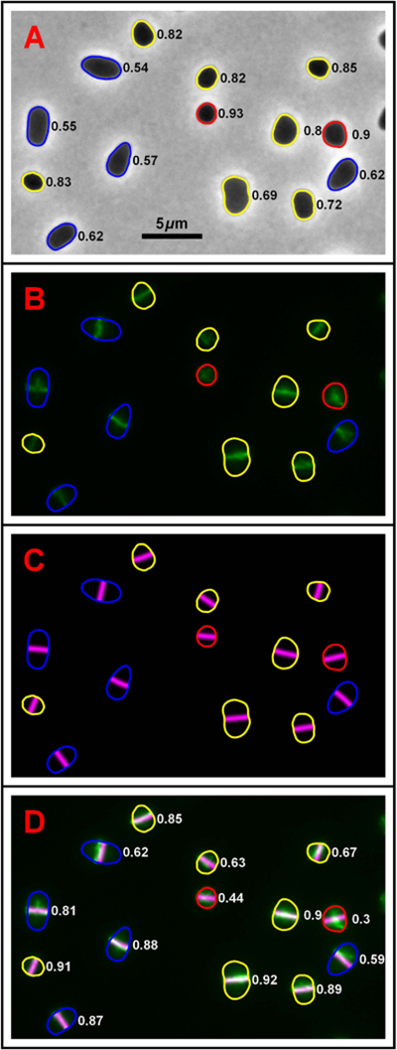
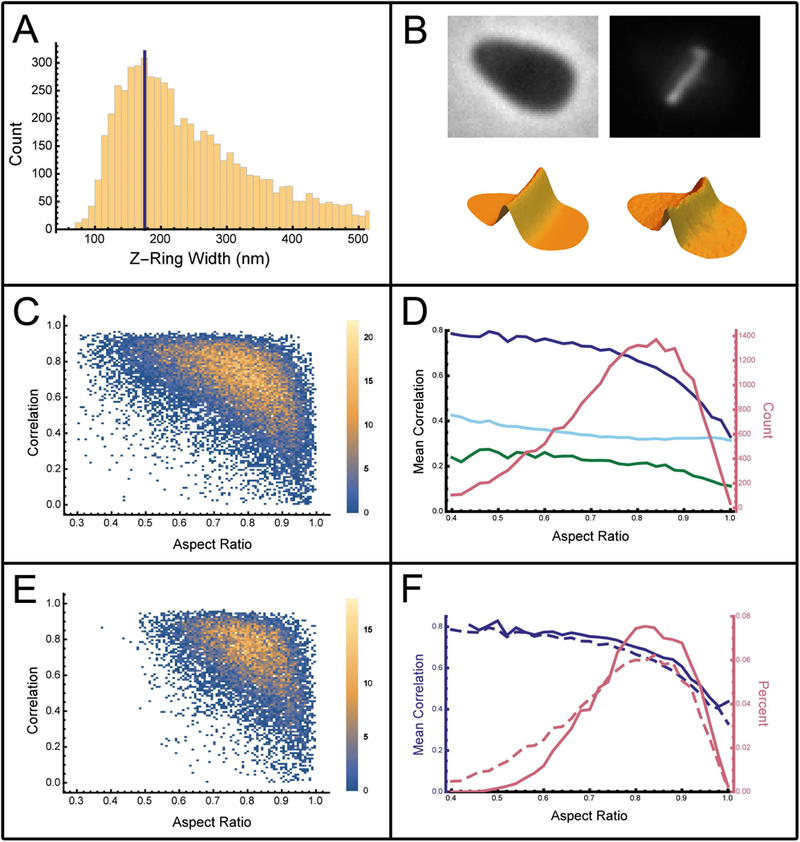
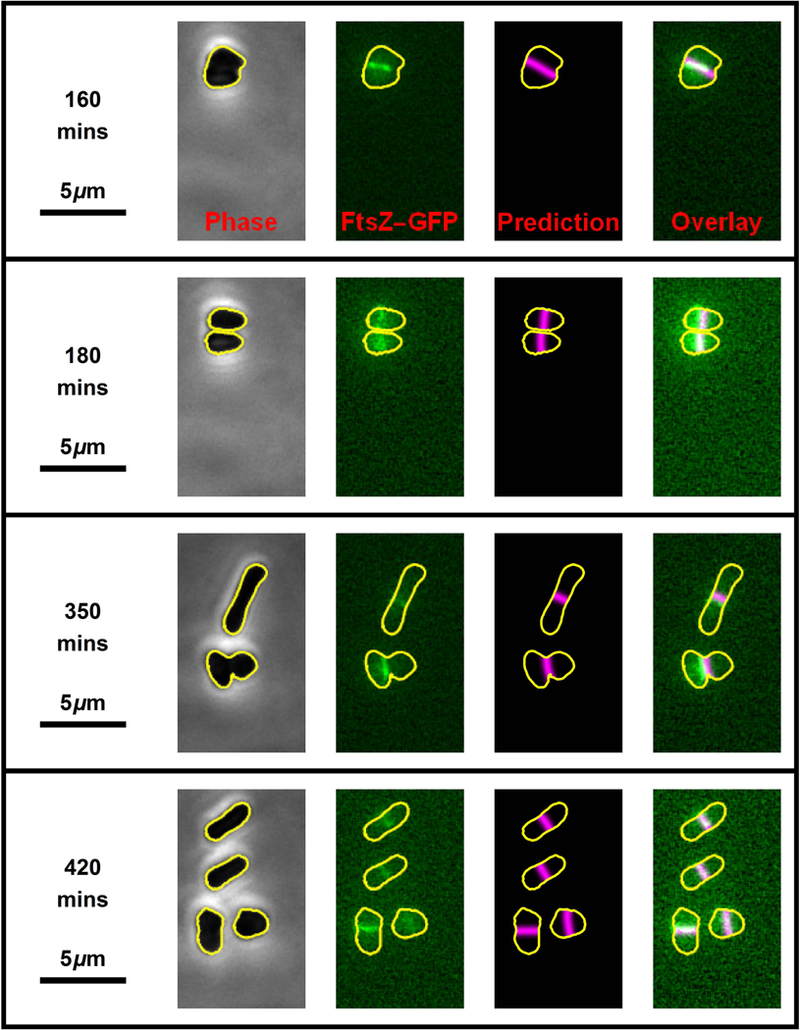
One mechanism for achieving accurate placement of the cell division machinery is via Turing patterns, where nonlinear molecular interactions spontaneously produce spatiotemporal concentration gradients. The resulting patterns are dictated by cell shape. For example, the Min system of Escherichia coli shows spatiotemporal oscillation between cell poles, leaving a mid-cell zone for division. The universality of pattern-forming mechanisms in divisome placement is currently unclear. We examined the location of the division plane in two pleomorphic archaea, Haloferax volcanii and Haloarcula japonica, and showed that it correlates with the predictions of Turing patterning. Time-lapse analysis of H. volcanii shows that divisome locations after successive rounds of division are dynamically determined by daughter cell shape. For H. volcanii, we show that the location of DNA does not influence division plane location, ruling out nucleoid occlusion. Triangular cells provide a stringent test for Turing patterning, where there is a bifurcation in division plane orientation. For the two archaea examined, most triangular cells divide as predicted by a Turing mechanism; however, in some cases multiple division planes are observed resulting in cells dividing into three viable progeny. Our results suggest that the division site placement is consistent with a Turing patterning system in these archaea.
© 2019 John Wiley & Sons Ltd.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
