

Binding to The Target Cell Surface Is The Crucial Step in Pore Formation of Hemolysin BL from Bacillus cereus
- PMID: 31137585
- PMCID: PMC6563250
- DOI: 10.3390/toxins11050281
Binding to The Target Cell Surface Is The Crucial Step in Pore Formation of Hemolysin BL from Bacillus cereus
Abstract
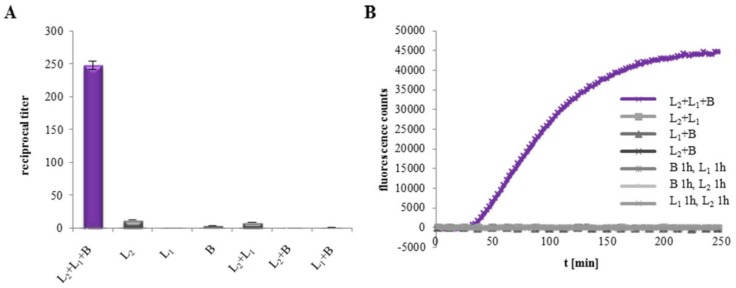
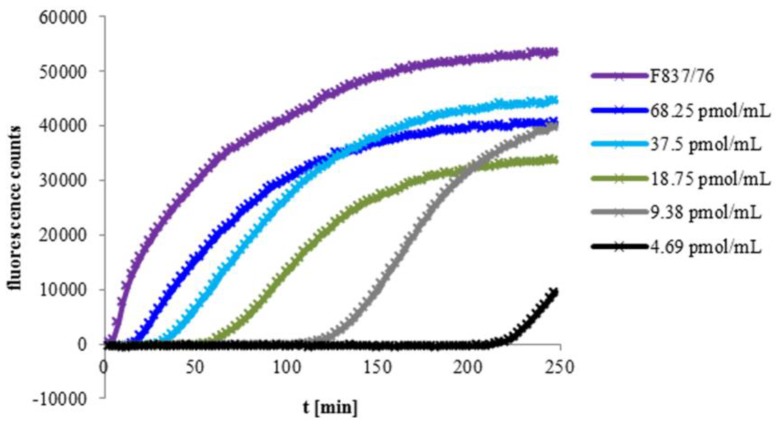
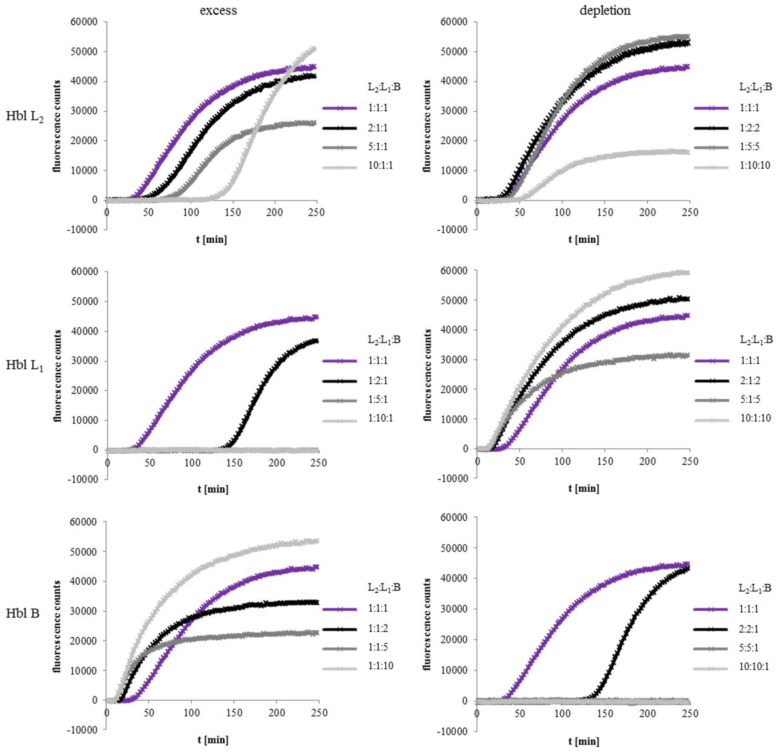
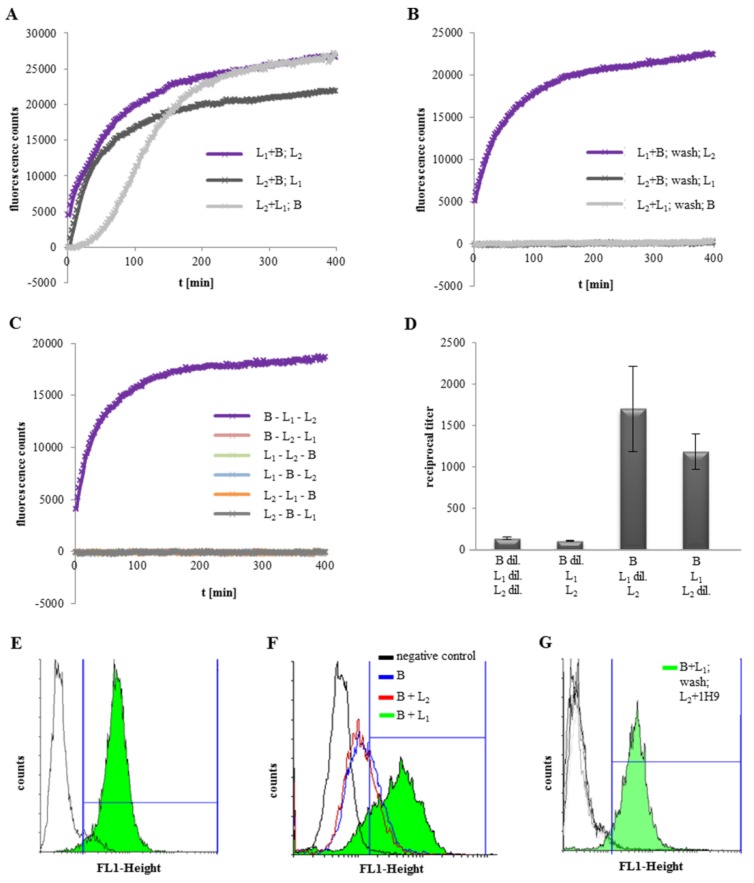
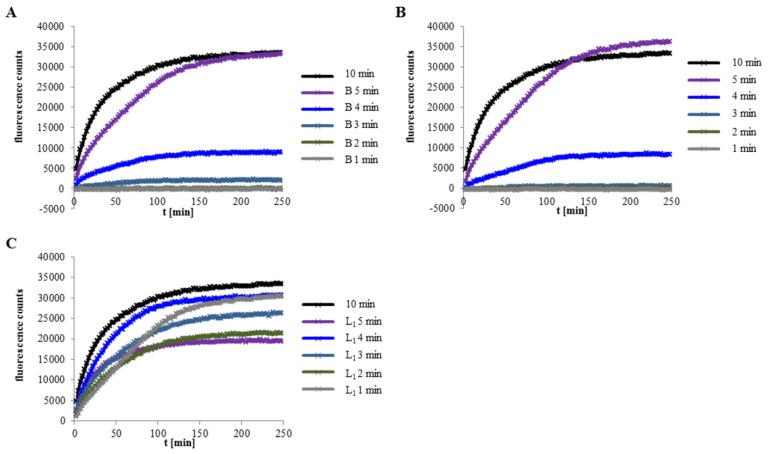
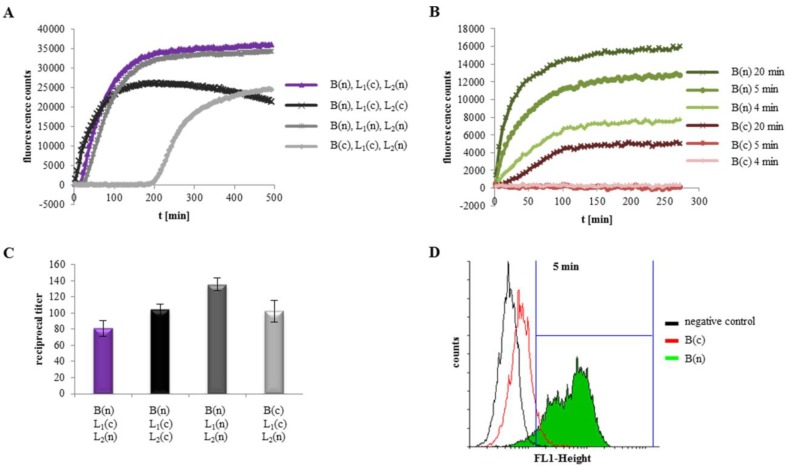
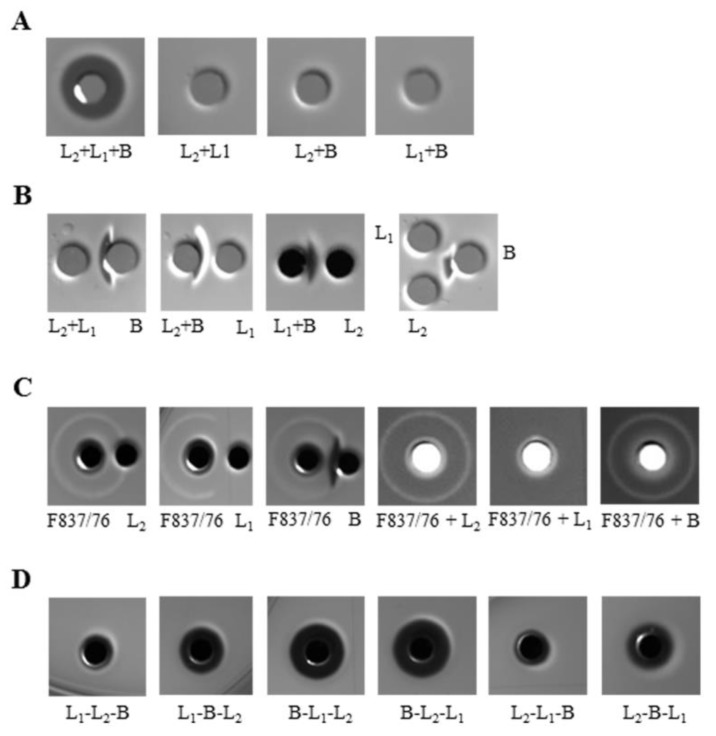
A major virulence factor involved in Bacillus cereus food poisoning is the three-component enterotoxin hemolysin BL. It consists of the binding component B and the two lytic components L1 and L2. Studying its mode of action has been challenging, as natural culture supernatants additionally contain Nhe, the second three-component enterotoxin, and purification of recombinant (r) Hbl components has been difficult. In this study, we report on pore-forming, cytotoxic, cell binding and hemolytic activity of recently generated rHbl components expressed in E. coli. It is known that all three Hbl components are necessary for cytotoxicity and pore formation. Here we show that an excess of rHbl B enhances, while an excess of rHbl L1 hinders, the velocity of pore formation. Most rapid pore formation was observed with ratios L2:L1:B = 1:1:10 and 10:1:10. It was further verified that Hbl activity is due to sequential binding of the components B - L1 - L2. Accordingly, all bioassays proved that binding of Hbl B to the cell surface is the crucial step for pore formation and cytotoxic activity. Binding of Hbl B took place within minutes, while apposition of the following L1 and L2 occurred immediately. Further on, applying toxin components simultaneously, it seemed that Hbl L1 enhanced binding of B to the target cell surface. Overall, these data contribute significantly to the elucidation of the mode of action of Hbl, and suggest that its mechanism of pore formation differs substantially from that of Nhe, although both enterotoxin complexes are sequentially highly related.
Keywords: Bacillus cereus; enterotoxins; hemolysin BL; mode of action; three-component toxin.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
-
- European Food Safety Authority The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks 2011. EFSA J. 2013;11:3129.
-
- Stenfors Arnesen L.P., Fagerlund A., Granum P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008;32:579–606. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
