

Transcriptome Response of Female Culicoides sonorensis Biting Midges (Diptera: Ceratopogonidae) to Early Infection with Epizootic Hemorrhagic Disease Virus (EHDV-2)
- PMID: 31137627
- PMCID: PMC6563219
- DOI: 10.3390/v11050473
Transcriptome Response of Female Culicoides sonorensis Biting Midges (Diptera: Ceratopogonidae) to Early Infection with Epizootic Hemorrhagic Disease Virus (EHDV-2)
Abstract
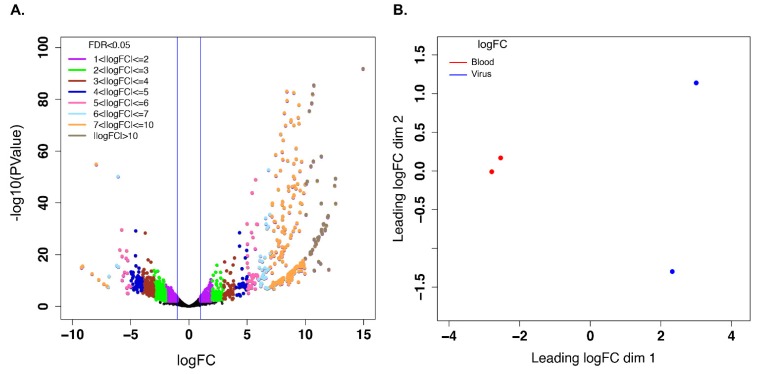
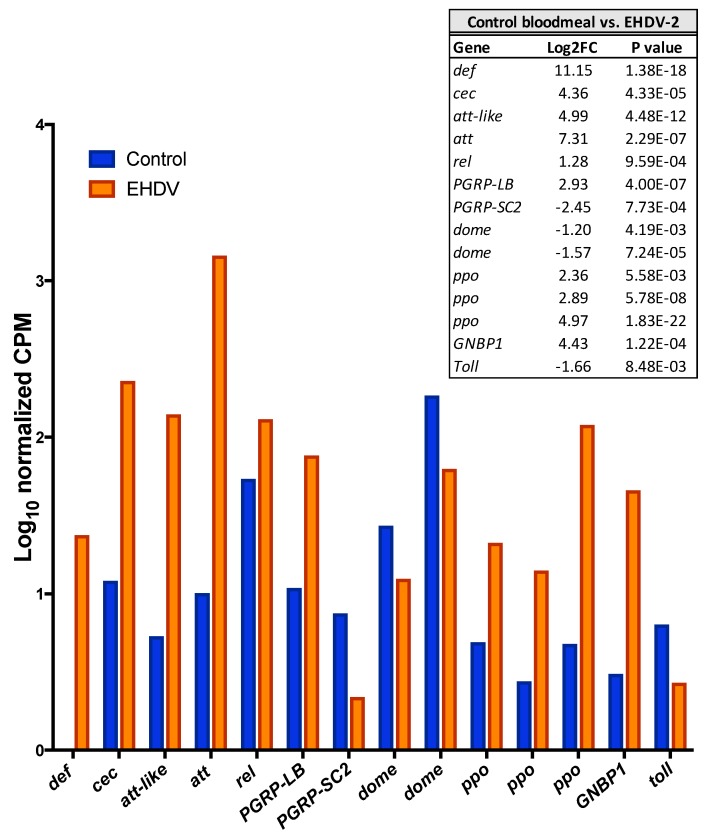
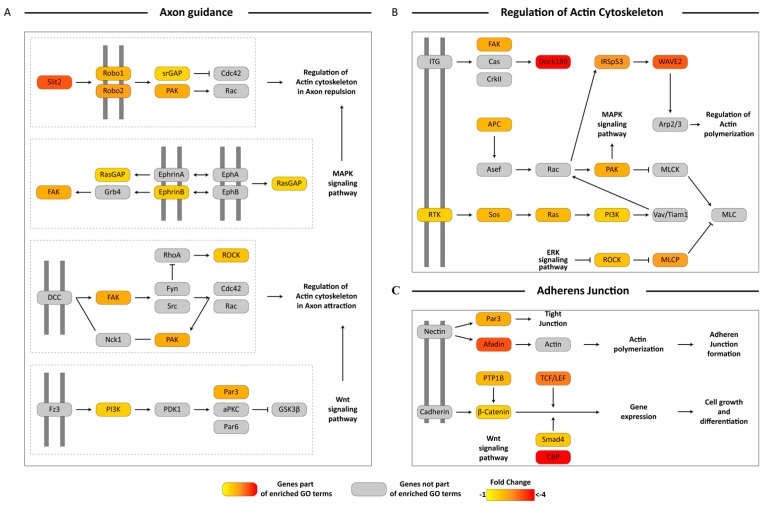
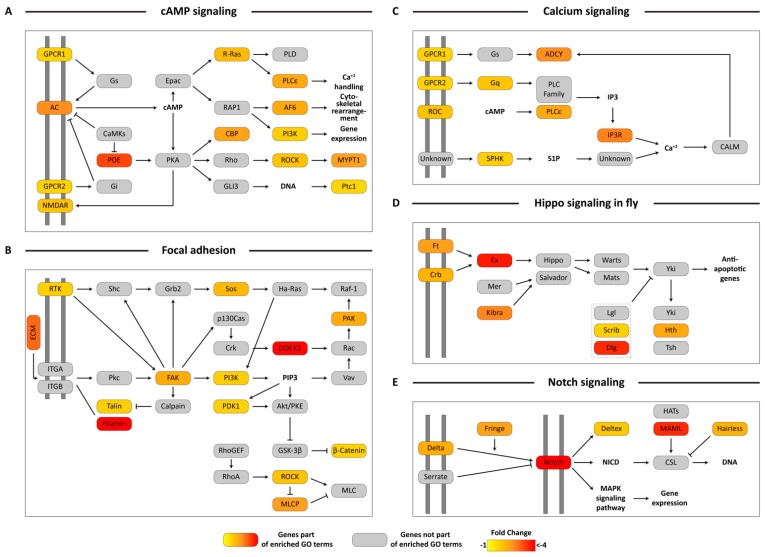
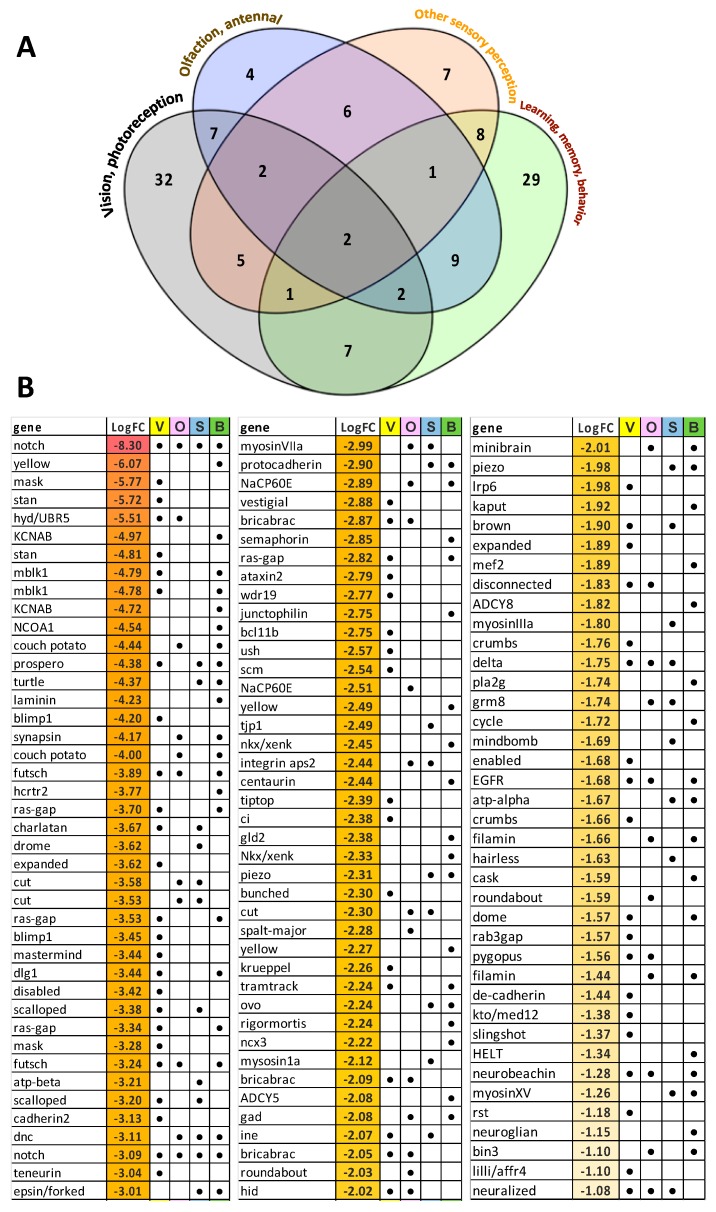
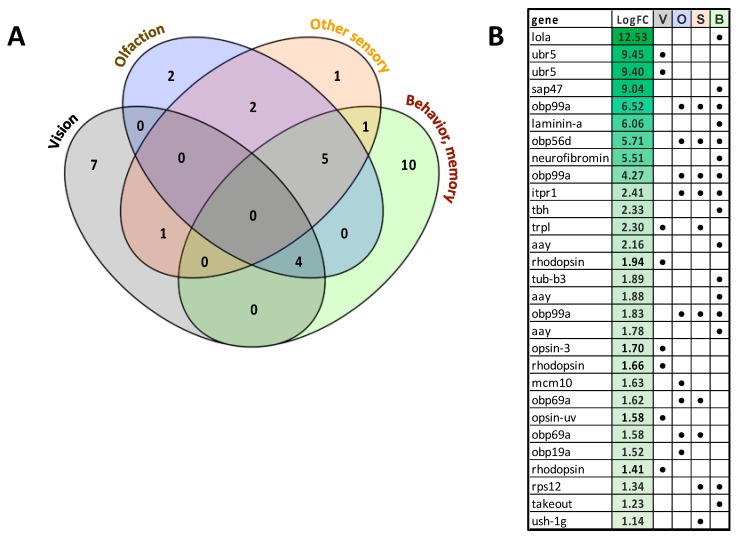
Female Culicoides sonorensis biting midges are vectors of epizootic hemorrhagic disease virus (EHDV), which causes morbidity and mortality in wild and domesticated ruminants. The aims in this study were to identify key changes in female midge transcriptome profiles occurring during early infection with EHDV-2. Midges were fed either negative control bloodmeals or bloodmeals containing EHDV-2 and transcriptomes were acquired at 36 h through deep sequencing. Reads were de novo assembled into a transcriptome comprised of 18,754 unigenes. Overall, there were 2401 differentially expressed unigenes and ~60% were downregulated in response to the virus (953 up; 1448 down). Downstream Gene Ontology enrichment, KEGG pathway mapping, and manual analyses were used to identify the effect of virus ingestion at both the gene and pathway levels. Downregulated unigenes were predominantly assigned to pathways related to cell/tissue structure and integrity (actin cytoskeleton, adherens junction, focal adhesion, hippo signaling), calcium signaling, eye morphogenesis and axon guidance. Unigenes attributed to sensory functions (especially vision), behavior, learning and memory were largely downregulated. Upregulated unigenes included those coding for innate immune processes, olfaction and photoreceptor pigments. Our results suggest that midges respond to virus infection as soon as 36 h post-ingestion, and that EHDV-2 may have a significant phenotypic effect on sensory and neural tissues.
Keywords: Culicoides; arbovirus; differential expression; dissemination; epizootic hemorrhagic disease virus; innate immune; midge; vector; visual perception.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ruder M.G., Lysyk T.J., Stallknecht D.E., Foil L.D., Johnson D.J., Chase C.C., Dargatz D.A., Gibbs E.P.J. Transmission and epidemiology of bluetongue and epizootic hemorrhagic disease in North America: Current perspectives, research gaps, and future directions. Vector-Borne Zoonotic Dis. 2015;15:348–363. doi: 10.1089/vbz.2014.1703. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
