

Serine/Arginine-Rich Splicing Factor 3 Modulates the Alternative Splicing of Cytoplasmic Polyadenylation Element Binding Protein 2
- PMID: 31138601
- PMCID: PMC6726571
- DOI: 10.1158/1541-7786.MCR-18-1291
Serine/Arginine-Rich Splicing Factor 3 Modulates the Alternative Splicing of Cytoplasmic Polyadenylation Element Binding Protein 2
Abstract
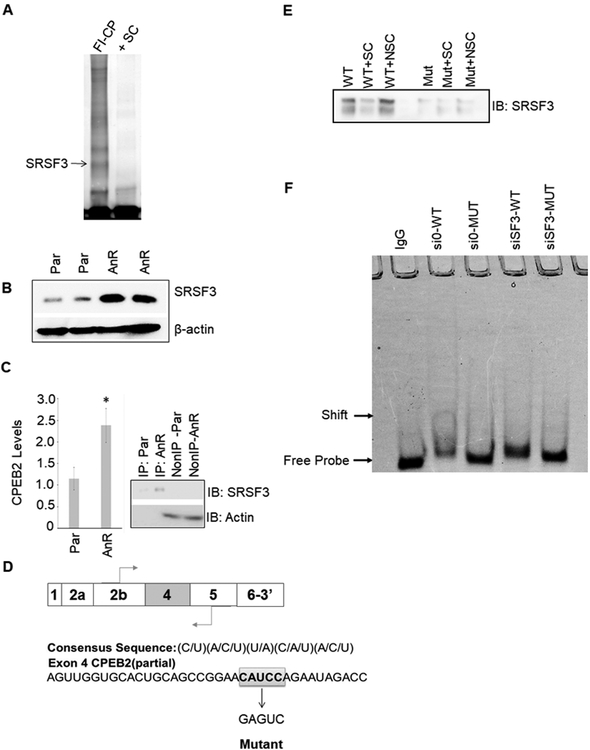
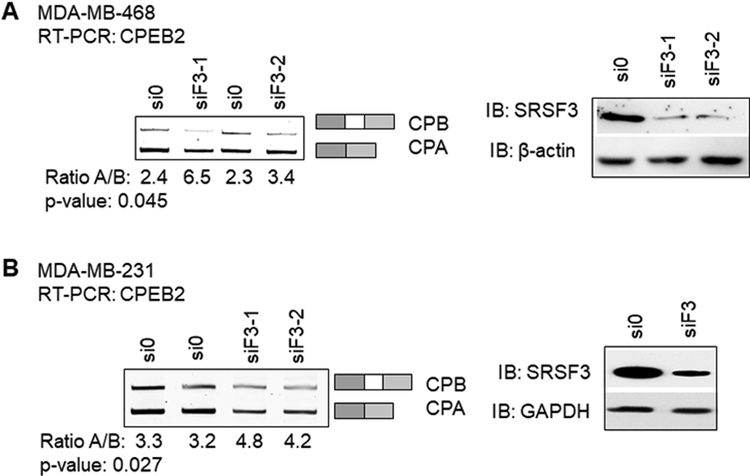
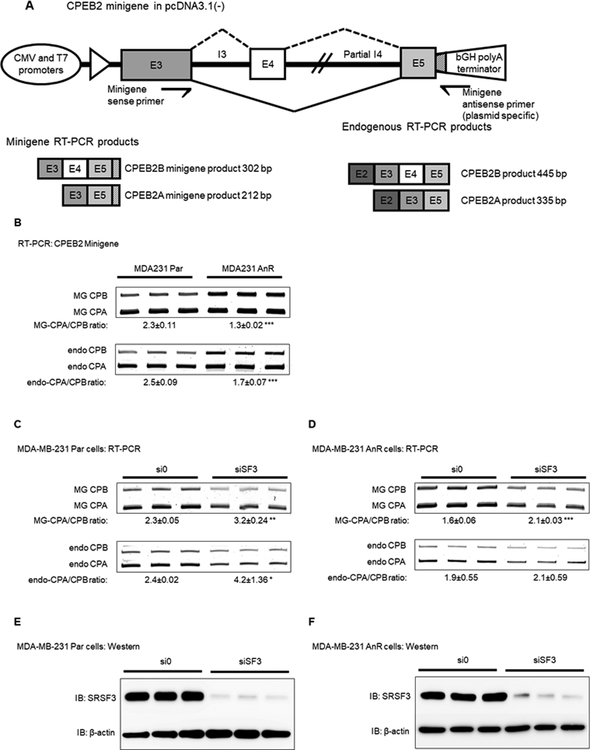
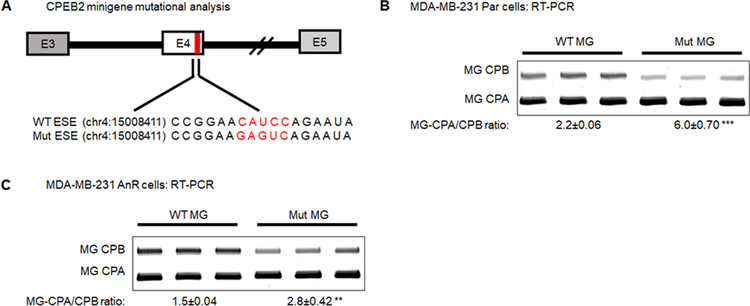
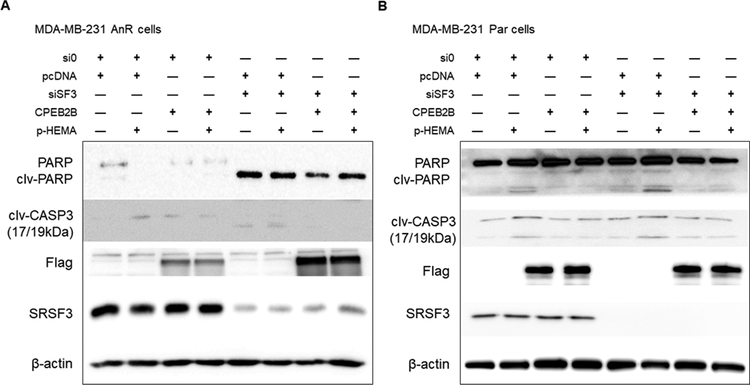
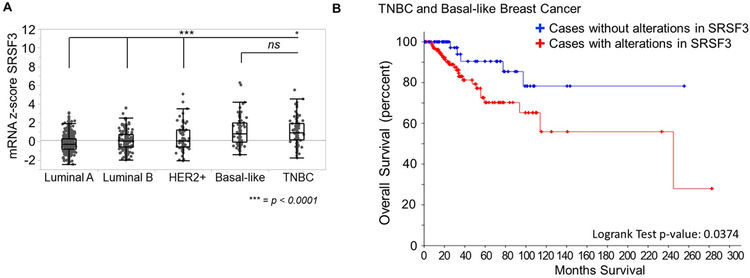
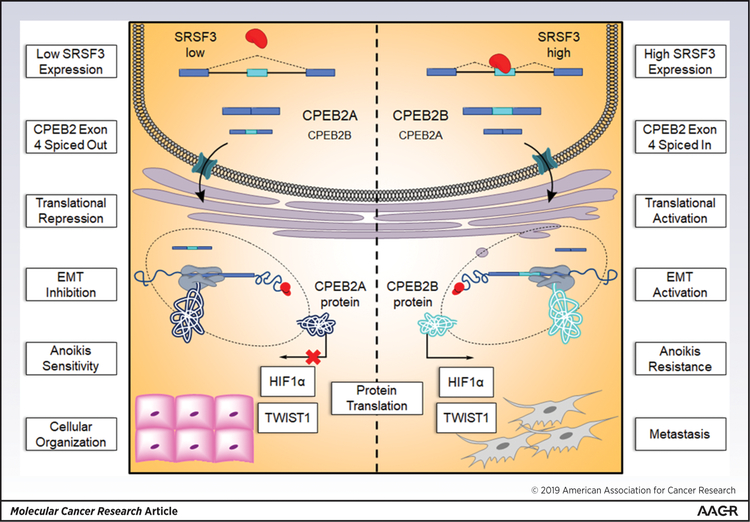
Triple negative breast cancer (TNBC) has an unusually low 5-year survival rate linked to higher metastatic rates. Our laboratory recently delineated a role for the alternative RNA splicing (AS) of cytoplasmic polyadenylation element binding protein 2 (CPEB2), via inclusion/exclusion of exon 4, in the metastasis of TNBC. In these studies, the mechanism governing the inclusion/exclusion of exon 4 was examined. Specifically, the RNA trans-factor, SRSF3, was found to be explicitly associated with CPEB2 exon 4. A SRSF3 consensus sequence was identified in exon 4, and mutation of this sequence abolished the association of SRSF3. The expression of SRSF3 was upregulated in TNBC cells upon the acquisition of anoikis resistance correlating with a reduction in the CPEB2A/B ratio. Importantly, downregulation of SRSF3 in these cells by siRNA induced the exclusion of exon 4 in cells increasing the ratio of CPEB2A (exon 4 excluded) to CPEB2B (exon 4 included). Downregulation of SRSF3 also reversed the CPEB2A/B ratio of a wild-type CPEB2 exon 4 minigene and endogenous CPEB2 pre-mRNA, but not a mutant CPEB2 minigene with the SRSF3 RNA cis-element ablated. SRSF3 downregulation ablated the anoikis resistance of TNBC cells, which was "rescued" by ectopic expression of CPEB2B. Finally, analysis of The Cancer Genome Atlas database showed a positive relationship between SRSF3 expression and lower CPEB2A/B ratios in aggressive breast cancers. IMPLICATIONS: These findings demonstrate that SRSF3 modulates CPEB2 AS to induce the expression of the CPEB2B isoform that drives TNBC phenotypes correlating with aggressive human breast cancer. VISUAL OVERVIEW: http://mcr.aacrjournals.org/content/molcanres/17/9/1920/F1.large.jpg.
©2019 American Association for Cancer Research.
Conflict of interest statement
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Schmadeka R, Harmon BE, and Singh M (2014) Triple-negative breast carcinoma: current and emerging concepts. Am J Clin Pathol 141, 462–477 - PubMed
-
- Dent R, Trudeau M, Pritchard KI, Hanna WM, Kahn HK, Sawka CA, Lickley LA, Rawlinson E, Sun P, and Narod SA (2007) Triple-negative breast cancer: clinical features and patterns of recurrence. Clin Cancer Res 13, 4429–4434 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
