

Translational downregulation of Twist1 expression by antiproliferative gene, B-cell translocation gene 2, in the triple negative breast cancer cells
- PMID: 31138781
- PMCID: PMC6538657
- DOI: 10.1038/s41419-019-1640-z
Translational downregulation of Twist1 expression by antiproliferative gene, B-cell translocation gene 2, in the triple negative breast cancer cells
Abstract
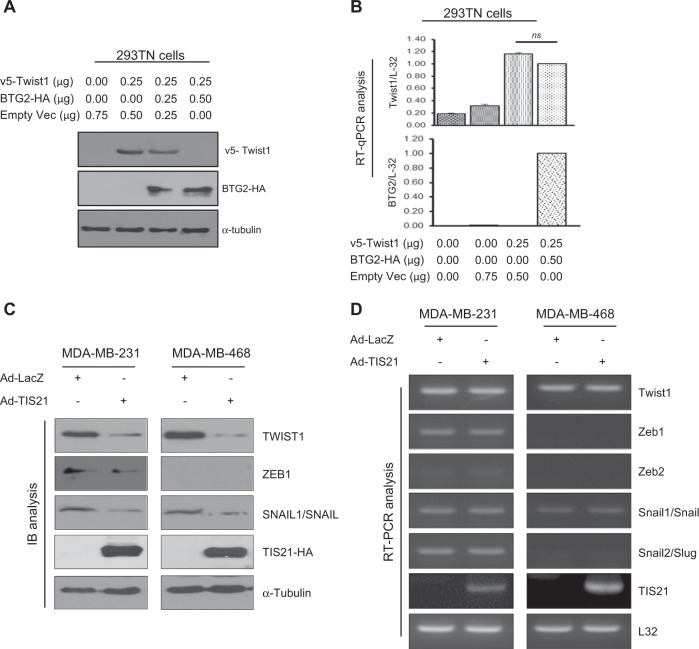
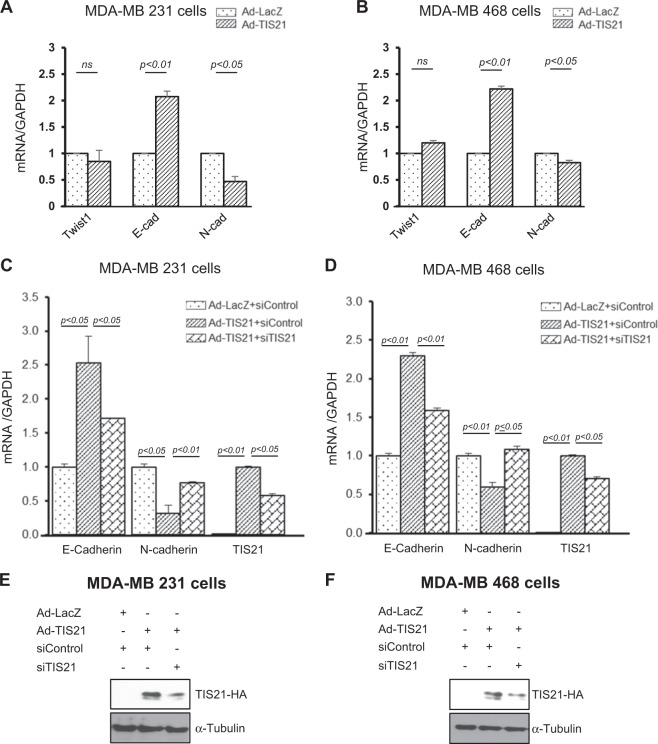
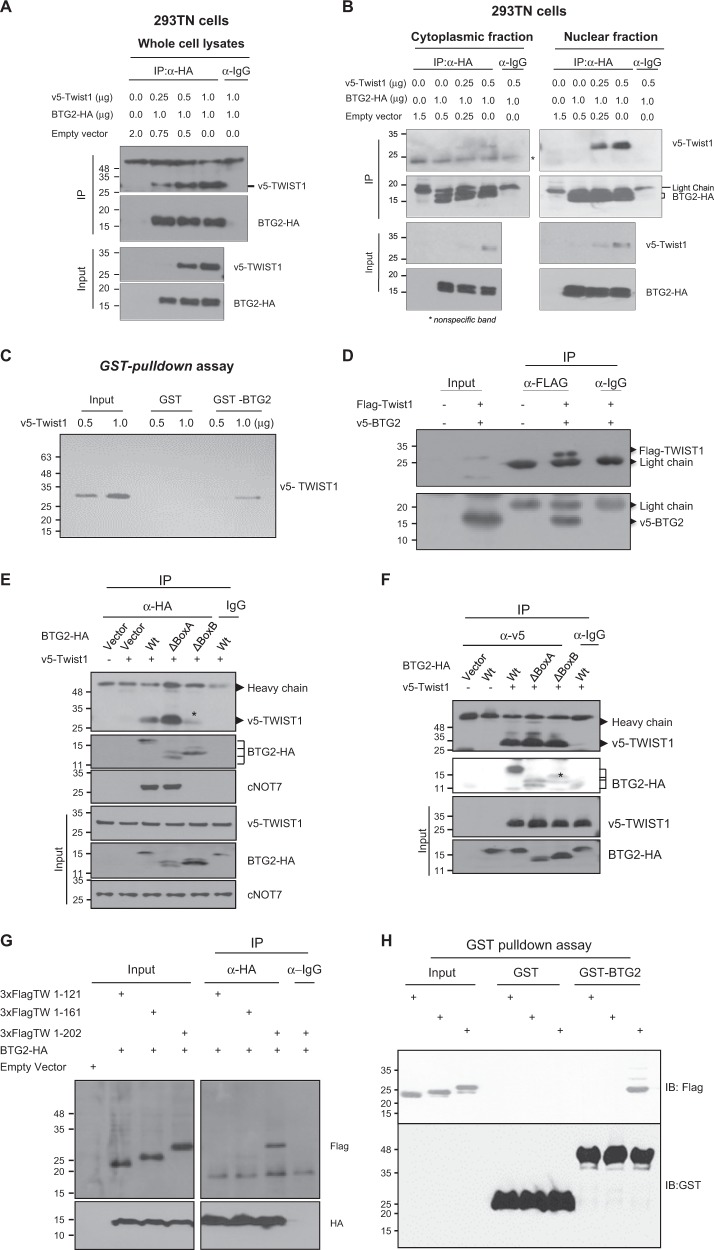
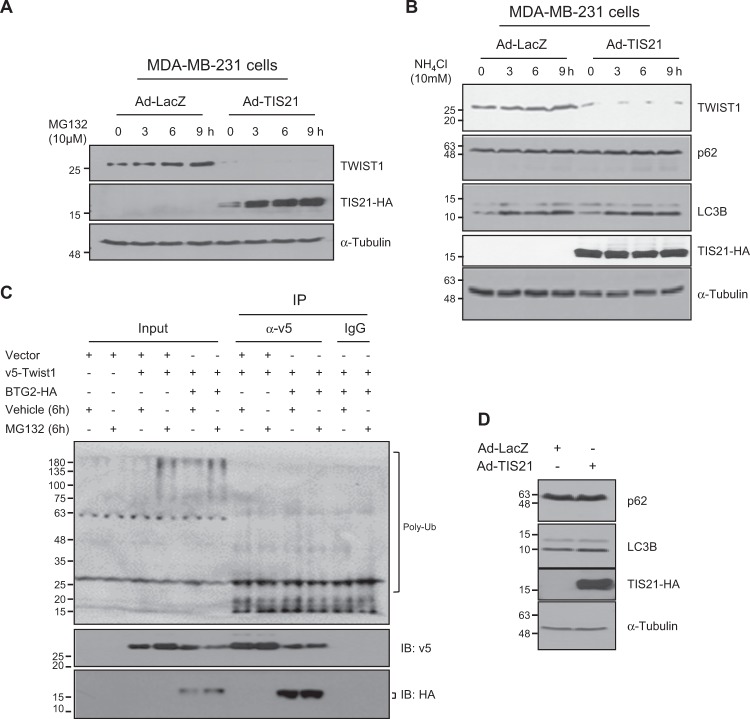
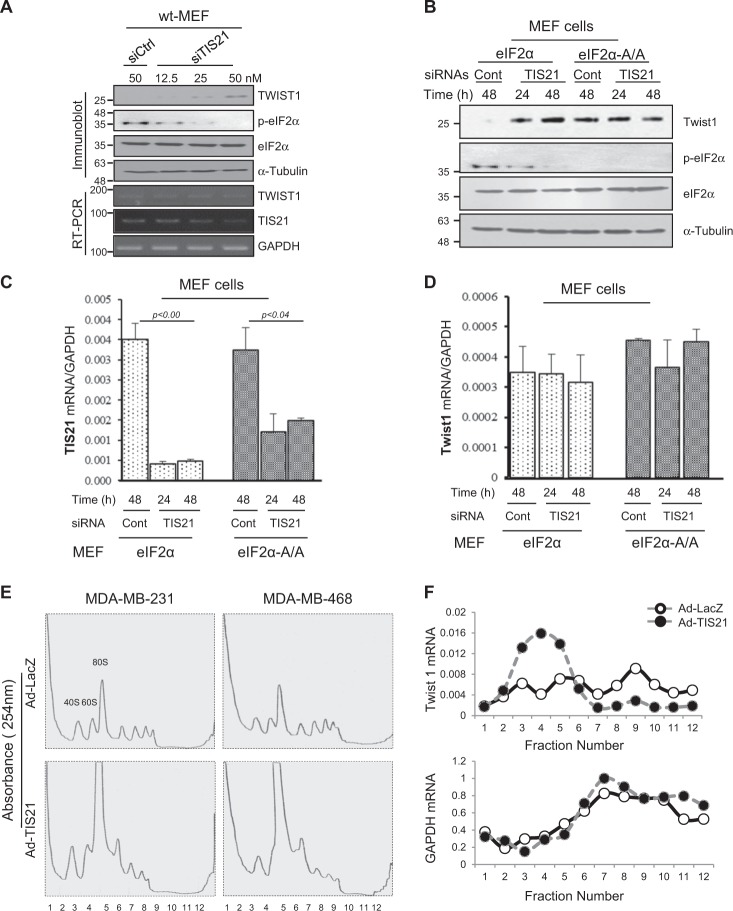
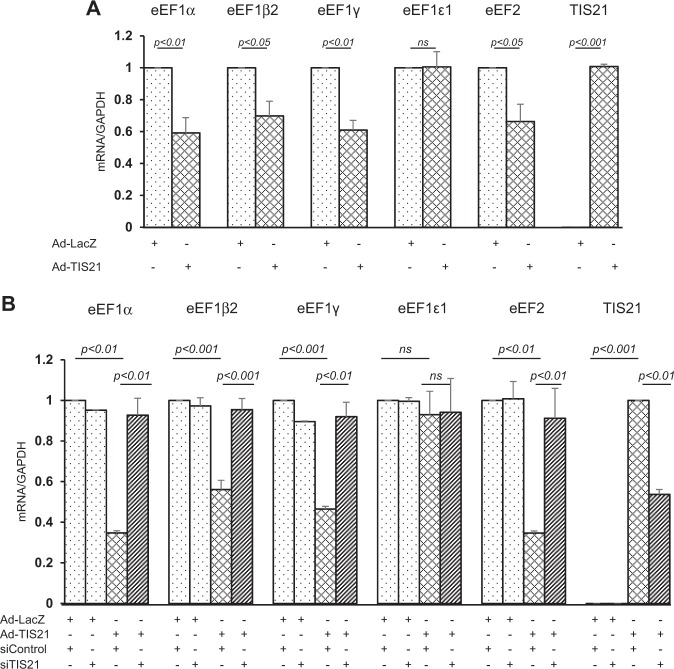
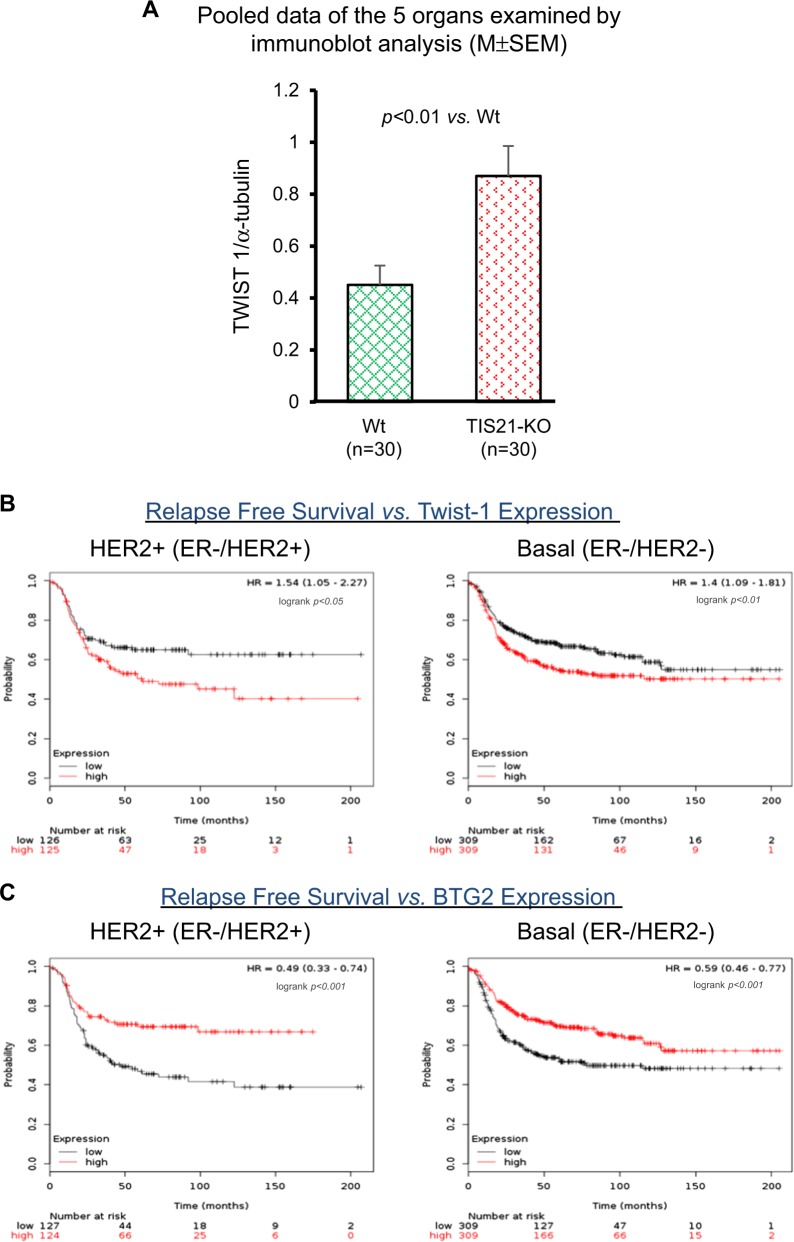
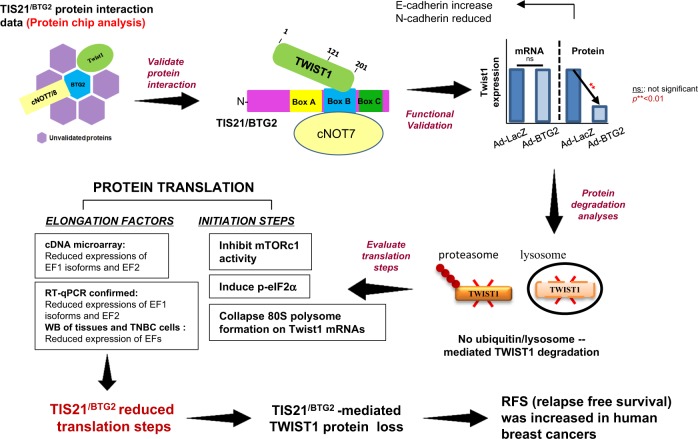
Twist1, a key transcription factor regulating epithelial-mesenchymal transition and cancer metastasis, is highly expressed in invasive cancers in contrast to the loss of BTG2/TIS21 expression. Based on our observation that forced expression of BTG2/TIS21 downregulated Twist1 protein expression without altering mRNA level, we investigated molecular mechanisms of the BTG2/TIS21-inhibited Twist1 translation in the triple negative breast cancer (TNBC) cells and in vivo BTG2/TIS21-knockout (KO) mice and human breast cancer tissues. (1) C-terminal domain of Twist1 and Box B of BTG2/TIS21 interacted with each other, which abrogated Twist1 activity. (2) BTG2/TIS21 inhibited translational initiation by depleting eIF4E availability via inhibiting 4EBP1 phosphorylation. (3) Expression of BTG2/TIS21 maintained p-eIF2α that downregulates initiation of protein translation, confirmed by eIF2α-AA mutant expression and BTG2/TIS21 knockdown in MEF cells. (4) cDNA microarray analysis revealed significantly higher expression of initiation factors-eIF2A, eIF3A, and eIF4G2-in the BTG2/TIS21-KO mouse than that in the wild type. (5) BTG2/TIS21-inhibited translation initiation lead to the collapse of polysome formation and the huge peak of 80s monomer in the BTG2/TIS21 expresser, but not in the control. (6) mRNAs and protein expressions of elongation factors were also downregulated by BTG2/TIS21 expression in TNBC cells, but much higher in both TIS21-KO mice and lymph node-positive human breast cancers. (7) BTG2/TIS21-mediated Twist1 loss was not due to the protein degradation by ubiquitination and autophagy activation. (8) Twist1 protein level was significantly higher in various organs of TIS21-KO mice compared with that in the control, indicating the in vivo role of BTG2/TIS21 gene in the regulation of Twist1 protein level. Altogether, the present study support our hypothesis that BTG2/TIS21 is a promising target to combat with metastatic cancers with high level of Twist1 without BTG2/TIS21 expression.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
